Vous aimerez peut-être aussi
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- FRQ Vocabulary ReviewDocument1 pageFRQ Vocabulary ReviewDrew AbbottPas encore d'évaluation
- 8 3ThingWorxInstallation 1 0 174675 PDFDocument157 pages8 3ThingWorxInstallation 1 0 174675 PDFAuditio MandhanyPas encore d'évaluation
- Lecture Planner - Chemistry - MANZIL For JEE 2024Document1 pageLecture Planner - Chemistry - MANZIL For JEE 2024Rishi NairPas encore d'évaluation
- STRESS HealthDocument40 pagesSTRESS HealthHajra KhanPas encore d'évaluation
- EUROJAM Diary3Document4 pagesEUROJAM Diary3Susan BakerPas encore d'évaluation
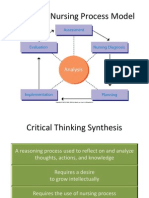
- NUR 104 Nursing Process MY NOTESDocument77 pagesNUR 104 Nursing Process MY NOTESmeanne073100% (1)
- Data Abstraction and Problem Solving With C Walls and Mirrors 6th Edition Carrano Solutions ManualDocument36 pagesData Abstraction and Problem Solving With C Walls and Mirrors 6th Edition Carrano Solutions Manualallocaturnonylgvtt100% (12)
- Soal PTS Vii BigDocument6 pagesSoal PTS Vii Bigdimas awe100% (1)
- BOMA - Office Building Standard Methods of Measurement AppendixDocument41 pagesBOMA - Office Building Standard Methods of Measurement Appendixxavest100% (7)
- ISA Guidelines PPF 1Document19 pagesISA Guidelines PPF 1Vasu DevanPas encore d'évaluation
- Lesson Plan Ordinal NumbersDocument5 pagesLesson Plan Ordinal Numbersapi-329663096Pas encore d'évaluation
- Turning The Mind Into An AllyDocument244 pagesTurning The Mind Into An AllyNic Sosa67% (3)
- A Technical Report ON Centre of Pressure ONA Plane Surface ExperimentDocument13 pagesA Technical Report ON Centre of Pressure ONA Plane Surface ExperimentVictor OwolekePas encore d'évaluation
- Proac Studio 100: Monitor Level Performance From An Established Compact DesignDocument2 pagesProac Studio 100: Monitor Level Performance From An Established Compact DesignAnonymous c3vuAsWAPas encore d'évaluation
- Spies May Be Gathering Encrypted Data To Crack With Future Quantum ComputerDocument1 pageSpies May Be Gathering Encrypted Data To Crack With Future Quantum ComputerHÀ ĐỖ VIẾTPas encore d'évaluation
- Marisa Wolf Final New ResumeDocument2 pagesMarisa Wolf Final New Resumeapi-403499166Pas encore d'évaluation
- Science: BiologyDocument22 pagesScience: BiologyMike RollidePas encore d'évaluation
- 5024Document2 pages5024Luis JesusPas encore d'évaluation
- VRF-SLB013-EN - 0805115 - Catalogo Ingles 2015 PDFDocument50 pagesVRF-SLB013-EN - 0805115 - Catalogo Ingles 2015 PDFJhon Lewis PinoPas encore d'évaluation
- Solution Manual-Statistical Physics of Particles by Meheran KardarDocument165 pagesSolution Manual-Statistical Physics of Particles by Meheran KardarDanielle Nguyen7% (14)
- LAB Find Out The Bottle ContentsDocument4 pagesLAB Find Out The Bottle ContentschanPas encore d'évaluation
- CHAPTER 2 (C) Innovation in EntrepreneurDocument36 pagesCHAPTER 2 (C) Innovation in EntrepreneurHuiLingPas encore d'évaluation
- HLA HART Concept of LawDocument19 pagesHLA HART Concept of LawHarneet KaurPas encore d'évaluation
- IO5 Future Skills Foresight 2030 ReportDocument96 pagesIO5 Future Skills Foresight 2030 ReportjuliavallePas encore d'évaluation
- NBCC 2015 Seismic Design Examples in S-FRAME AnalysisDocument91 pagesNBCC 2015 Seismic Design Examples in S-FRAME AnalysisMike Smith100% (1)
- Ielts Reading Actual Tests With Suggested Answers Oct 2021 JDocument508 pagesIelts Reading Actual Tests With Suggested Answers Oct 2021 JHarpreet Singh JohalPas encore d'évaluation
- Automotive SensorsDocument20 pagesAutomotive SensorsahmedPas encore d'évaluation
- Review of Ventura's "An Overview of Child Psychology in The Philippines"Document2 pagesReview of Ventura's "An Overview of Child Psychology in The Philippines"Irene CayePas encore d'évaluation
- Lesson 7Document6 pagesLesson 7Jeya Plays YTPas encore d'évaluation
- Test Bank For Body Structures and Functions 11th Edition Ann ScottDocument27 pagesTest Bank For Body Structures and Functions 11th Edition Ann Scottpellucidutter.uka6lr100% (38)