Vous aimerez peut-être aussi
- Human ScleraDocument15 pagesHuman ScleraSAMPas encore d'évaluation
- SD CohenDocument8 pagesSD CohenManuel VasquezPas encore d'évaluation
- Glaucoma A Symposium Presented at a Meeting of the Chicago Ophthalmological Society, November 17, 1913D'EverandGlaucoma A Symposium Presented at a Meeting of the Chicago Ophthalmological Society, November 17, 1913Pas encore d'évaluation
- Corneal Biomechanical Changes and Intraocular Pressure in Patients With Thyroid OrbitopathyDocument5 pagesCorneal Biomechanical Changes and Intraocular Pressure in Patients With Thyroid Orbitopathykevin_jawanPas encore d'évaluation
- Comparison of Corneal Biomechanical Properties Among AxialDocument8 pagesComparison of Corneal Biomechanical Properties Among AxialPutri kartiniPas encore d'évaluation
- Blood Pressure and GlaucomaDocument6 pagesBlood Pressure and GlaucomaRizqy AbdullahPas encore d'évaluation
- JCI61135Document16 pagesJCI61135Cahya Daris TriwibowoPas encore d'évaluation
- Handbook of Clinical Electrophysiology of VisionD'EverandHandbook of Clinical Electrophysiology of VisionMinzhong YuPas encore d'évaluation
- High Myopia and Glaucoma Susceptibility The Beijing Eye StudyDocument5 pagesHigh Myopia and Glaucoma Susceptibility The Beijing Eye StudyDickyStevanoZukhriPas encore d'évaluation
- 2012 Cer Hiraoka Long-Term Effect of Orthokeratology 5yDocument7 pages2012 Cer Hiraoka Long-Term Effect of Orthokeratology 5yIgnacio AlvarezPas encore d'évaluation
- Inflammation and The Nervous System: The Connection in The Cornea in Patients With Infectious KeratitisDocument8 pagesInflammation and The Nervous System: The Connection in The Cornea in Patients With Infectious KeratitisrizkydimasaputraPas encore d'évaluation
- Human Optical Axial Length and DefocusDocument8 pagesHuman Optical Axial Length and DefocusvchengsongPas encore d'évaluation
- Stem Cell Biology and Regenerative Medicine in OphthalmologyD'EverandStem Cell Biology and Regenerative Medicine in OphthalmologyStephen TsangPas encore d'évaluation
- DR RatnaDocument6 pagesDR RatnaNi Kadek Eno YuliantariPas encore d'évaluation
- GlaucomaDocument9 pagesGlaucomaREGINE YEO ZHI SHUENPas encore d'évaluation
- Medicina 59 01660 v2Document10 pagesMedicina 59 01660 v2Maria CorbuPas encore d'évaluation
- The Ocular Biometric and Corneal Topographic Characteristics of High-Anisometropic Adults in TaiwanDocument6 pagesThe Ocular Biometric and Corneal Topographic Characteristics of High-Anisometropic Adults in TaiwanNurul Falah KalokoPas encore d'évaluation
- Disorders of the Patellofemoral Joint: Diagnosis and ManagementD'EverandDisorders of the Patellofemoral Joint: Diagnosis and ManagementPas encore d'évaluation
- Characterizing Osteochondrosis in The Dog: Potential Roles For Matrix Metalloproteinases and Mechanical Load in Pathogenesis and Disease ProgressionDocument10 pagesCharacterizing Osteochondrosis in The Dog: Potential Roles For Matrix Metalloproteinases and Mechanical Load in Pathogenesis and Disease ProgressionSolange SantanaPas encore d'évaluation
- Rajiv Gandhi University of Health Sciences, Karnataka, Bangalore Annexure-Ii Proforma For Registration of Subject For DissertationDocument9 pagesRajiv Gandhi University of Health Sciences, Karnataka, Bangalore Annexure-Ii Proforma For Registration of Subject For DissertationNavin ChandarPas encore d'évaluation
- Risk Factors For Development of Phacomorphic Glaucoma in Mature Cataract - A Hospital Based Study, JhansiDocument5 pagesRisk Factors For Development of Phacomorphic Glaucoma in Mature Cataract - A Hospital Based Study, JhansiIJAR JOURNALPas encore d'évaluation
- TMP 82 D3Document5 pagesTMP 82 D3FrontiersPas encore d'évaluation
- Korneal HysteresisDocument5 pagesKorneal HysteresisIndana ZfPas encore d'évaluation
- Primary Open-Angle Glaucoma: SeminarDocument10 pagesPrimary Open-Angle Glaucoma: SeminarFebry LuthunananaPas encore d'évaluation
- Dynamic Scheimpflug Ocular Biomechanical Parameters in Healthy and Medically Controlled Glaucoma EyesDocument5 pagesDynamic Scheimpflug Ocular Biomechanical Parameters in Healthy and Medically Controlled Glaucoma EyesNetflix DheaPas encore d'évaluation
- Management of Paediatric Traumatic Cataract by Epilenticular Intraocular Lens Implantation: Long-Term Visual Results and Postoperative ComplicationsDocument5 pagesManagement of Paediatric Traumatic Cataract by Epilenticular Intraocular Lens Implantation: Long-Term Visual Results and Postoperative ComplicationsWahyu Tri UtomoPas encore d'évaluation
- Ocular Biometry in Patients With Primary Open Angle Glaucoma PoagDocument3 pagesOcular Biometry in Patients With Primary Open Angle Glaucoma Poagnabila tiara SantosoPas encore d'évaluation
- Wong 2003 Refractive Errors, Axial Ocular Dimensions, and Age-Related Cataracts The Tanjong Pagar SurveyDocument7 pagesWong 2003 Refractive Errors, Axial Ocular Dimensions, and Age-Related Cataracts The Tanjong Pagar SurveyYung Chun LinPas encore d'évaluation
- Jia Et Al. - 2016 - Biomechanics of The Sclera and Effects On Intraocu PDFDocument8 pagesJia Et Al. - 2016 - Biomechanics of The Sclera and Effects On Intraocu PDFFelicia SutarliPas encore d'évaluation
- Differences in Structural Parameters in Patients With Open-Angle Glaucoma, High Myopia and Both Diseases Concurrently. A Pilot StudyDocument14 pagesDifferences in Structural Parameters in Patients With Open-Angle Glaucoma, High Myopia and Both Diseases Concurrently. A Pilot Study钟华Pas encore d'évaluation
- Glaucoma - State of The Art and Perspectives On TreatmentDocument17 pagesGlaucoma - State of The Art and Perspectives On TreatmentbarbaraPas encore d'évaluation
- Preprints202303 0018 v1Document12 pagesPreprints202303 0018 v1vergarajaviera548Pas encore d'évaluation
- Link Between Neurodegeneration and Trabecular Meshwork Injury in Glaucomatous PatientsDocument7 pagesLink Between Neurodegeneration and Trabecular Meshwork Injury in Glaucomatous Patientsfaruq azmiPas encore d'évaluation
- Puccinelli 2019Document8 pagesPuccinelli 2019RathavishwarajPas encore d'évaluation
- Brandt 2001Document10 pagesBrandt 2001riyad saksakPas encore d'évaluation
- Prospective Evaluation of Acupuncture As Treatment For GlaucomaDocument10 pagesProspective Evaluation of Acupuncture As Treatment For Glaucoma16.11Hz MusicPas encore d'évaluation
- AmblyopiaDocument10 pagesAmblyopiaNoura RosePas encore d'évaluation
- Oct 3Document15 pagesOct 3Star LadyPas encore d'évaluation
- An Eye To Treating Blindness: News & ViewsDocument3 pagesAn Eye To Treating Blindness: News & ViewsEry Noer WibowoPas encore d'évaluation
- Effects of Primary and Recurrent Sacral Chordoma On The Motor and Nociceptive Function of Hindlimbs in Rats - An Orthotopic Spine ModelDocument12 pagesEffects of Primary and Recurrent Sacral Chordoma On The Motor and Nociceptive Function of Hindlimbs in Rats - An Orthotopic Spine ModelgangligonPas encore d'évaluation
- Efficacy and Toxicity of Intravitreous Chemotherapy For Retinoblastoma: Four-Year ExperienceDocument8 pagesEfficacy and Toxicity of Intravitreous Chemotherapy For Retinoblastoma: Four-Year ExperienceSlr RandiPas encore d'évaluation
- High Correlation of Scotopic and Photopic Electroretinogram Components With Severity of Central Retinal Artery OcclusionDocument7 pagesHigh Correlation of Scotopic and Photopic Electroretinogram Components With Severity of Central Retinal Artery OcclusionrosaPas encore d'évaluation
- tmp6236 TMPDocument12 pagestmp6236 TMPFrontiersPas encore d'évaluation
- Physiology PharmacologyDocument126 pagesPhysiology PharmacologyuneedlesPas encore d'évaluation
- Jurnal New3Document9 pagesJurnal New3Nurul Qomariah TPas encore d'évaluation
- Chance 2Document12 pagesChance 2Yahir Gamaliel Nava HidalgoPas encore d'évaluation
- BR J Ophthalmol-2014-Chan-79-81 PDFDocument4 pagesBR J Ophthalmol-2014-Chan-79-81 PDFRomi Mauliza FauziPas encore d'évaluation
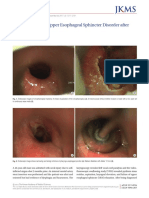
- Dysphagia Due To Upper Esophageal Sphincter DisordDocument3 pagesDysphagia Due To Upper Esophageal Sphincter DisordEustakia Rini Kartika DewiPas encore d'évaluation
- Steriod Cataract PDFDocument4 pagesSteriod Cataract PDFLisa IskandarPas encore d'évaluation
- Retinal Prosthesis: A Clinical Guide to Successful ImplementationD'EverandRetinal Prosthesis: A Clinical Guide to Successful ImplementationMark S. HumayunPas encore d'évaluation
- Morphology of The Murine Optic Nerve: Chr. Albrecht May and Elke Lu Tjen-DrecollDocument7 pagesMorphology of The Murine Optic Nerve: Chr. Albrecht May and Elke Lu Tjen-DrecollEzra MargarethPas encore d'évaluation
- Defocus Press OnDocument10 pagesDefocus Press OnLuigi PolitoPas encore d'évaluation
- Ceo0042 0665Document10 pagesCeo0042 0665walid abdallahPas encore d'évaluation
- JClinOphthalmolRes43123-8318319 021838Document4 pagesJClinOphthalmolRes43123-8318319 021838Dex Bayu AcPas encore d'évaluation
- Red Light MiopiaDocument11 pagesRed Light MiopiaLuigi PolitoPas encore d'évaluation
- Pseudophakic BullousDocument6 pagesPseudophakic BullousanglingPas encore d'évaluation
- Anorexia ReprintDocument6 pagesAnorexia ReprintdoctorwhoiswhoPas encore d'évaluation
- Glaucoma: A Disease of Early Cellular Senescence: Chrisye Lena Baura 2008-83-033Document20 pagesGlaucoma: A Disease of Early Cellular Senescence: Chrisye Lena Baura 2008-83-033Chrisye LeenaPas encore d'évaluation
- PIIS0002939422003774Document10 pagesPIIS0002939422003774Anca Florina GaceaPas encore d'évaluation
- DBD Grade IIDocument1 pageDBD Grade IIAtunk RahmanPas encore d'évaluation
- Hematology Disease in Geriatric (25!01!2016)Document1 pageHematology Disease in Geriatric (25!01!2016)Atunk RahmanPas encore d'évaluation
- Osteoid OsteomaDocument1 pageOsteoid OsteomaAtunk RahmanPas encore d'évaluation
- Fibroid KondromixoidDocument1 pageFibroid KondromixoidAtunk RahmanPas encore d'évaluation
- Hernia UmbilicalDocument1 pageHernia UmbilicalAtunk RahmanPas encore d'évaluation
- WHS2010 Part2Document103 pagesWHS2010 Part2Atunk RahmanPas encore d'évaluation
- DicDocument1 pageDicAtunk RahmanPas encore d'évaluation
- GTG 27Document26 pagesGTG 27Atunk RahmanPas encore d'évaluation
- Basic Surgical SkilDocument1 pageBasic Surgical SkilAtunk RahmanPas encore d'évaluation
- Meniere DiseaseDocument1 pageMeniere DiseaseAtunk RahmanPas encore d'évaluation
- GlaukomaDocument16 pagesGlaukomaAtunk RahmanPas encore d'évaluation
- BatukDocument28 pagesBatukAtunk RahmanPas encore d'évaluation
- Tension HeadacheDocument1 pageTension HeadacheAtunk RahmanPas encore d'évaluation
- Langkah Diagnosis EncephalitisDocument1 pageLangkah Diagnosis EncephalitisAtunk RahmanPas encore d'évaluation
- Cover Klipink TinkomDocument1 pageCover Klipink TinkomAtunk RahmanPas encore d'évaluation
- Colonic PolypDocument1 pageColonic PolypAtunk RahmanPas encore d'évaluation
- Radial Tunnel SyndromDocument1 pageRadial Tunnel SyndromAtunk RahmanPas encore d'évaluation
- Sch3u Exam Review Ws s2018 PDFDocument4 pagesSch3u Exam Review Ws s2018 PDFwdsfPas encore d'évaluation
- (Eng) Zx890lch 5a Ks En316Document13 pages(Eng) Zx890lch 5a Ks En316MC TAK LEEPas encore d'évaluation
- Efficient Rice Based Cropping SystemDocument24 pagesEfficient Rice Based Cropping Systemsenthilnathan100% (1)
- Bahir Dar University BIT: Faculity of Mechanical and Industrial EngineeringDocument13 pagesBahir Dar University BIT: Faculity of Mechanical and Industrial Engineeringfraol girmaPas encore d'évaluation
- Donna Hay Magazine 2014-10-11 PDFDocument172 pagesDonna Hay Magazine 2014-10-11 PDFlekovic_tanjaPas encore d'évaluation
- Tsel Manual ThermoselDocument25 pagesTsel Manual ThermoselPedro AlvelaisPas encore d'évaluation
- Banachek UnlimitedDocument34 pagesBanachek UnlimitedserenaPas encore d'évaluation
- Single Door Feeder Pillar 200A MCCBDocument1 pageSingle Door Feeder Pillar 200A MCCBMiqdad AliPas encore d'évaluation
- Math 202 Syllabus-Spring18 PDFDocument2 pagesMath 202 Syllabus-Spring18 PDFvonacoc49Pas encore d'évaluation
- Earth Science (Metamorphic Rocks)Document8 pagesEarth Science (Metamorphic Rocks)MA. ALEXIS LAURENPas encore d'évaluation
- Unit 2 - Presentations (Image, Impact and Making An Impression) 2Document25 pagesUnit 2 - Presentations (Image, Impact and Making An Impression) 2LK Chiarra Panaligan100% (1)
- Intumescent Paint For Steel NZ - Coating - Co.nzDocument8 pagesIntumescent Paint For Steel NZ - Coating - Co.nzPeter ThomsonPas encore d'évaluation
- AD1 Elements of Architectural Composition For Placemaking PDFDocument4 pagesAD1 Elements of Architectural Composition For Placemaking PDFAndre Yap100% (1)
- Mathematical Modeling of Mechanical Systems and Electrical SystemsDocument49 pagesMathematical Modeling of Mechanical Systems and Electrical SystemsMary DunhamPas encore d'évaluation
- Accsap 10 VHDDocument94 pagesAccsap 10 VHDMuhammad Javed Gaba100% (2)
- Book BindingDocument14 pagesBook Bindingpesticu100% (2)
- Spectroscopy (L-26 To 29)Document81 pagesSpectroscopy (L-26 To 29)Vashistha GargPas encore d'évaluation
- ODA X9 2SL DatasheetDocument8 pagesODA X9 2SL DatasheetYudy UtamaPas encore d'évaluation
- Astm 2017-F2059Document5 pagesAstm 2017-F2059laythPas encore d'évaluation
- Digital Signal Processing: B.E Ece (5Th Semester)Document17 pagesDigital Signal Processing: B.E Ece (5Th Semester)Saatwat CoolPas encore d'évaluation
- Rankine-Hugoniot Curve: CJ: Chapman JouguetDocument6 pagesRankine-Hugoniot Curve: CJ: Chapman Jouguetrattan5Pas encore d'évaluation
- Cell Structure, Function Practice Test With AnswersDocument16 pagesCell Structure, Function Practice Test With AnswersDJ ISAACSPas encore d'évaluation
- Eliminating Oscillation Between Parallel MnosfetsDocument6 pagesEliminating Oscillation Between Parallel MnosfetsCiprian BirisPas encore d'évaluation
- Chemical Bonds WorksheetDocument2 pagesChemical Bonds WorksheetJewel Mae MercadoPas encore d'évaluation
- Natural Disasters Vocabulary Exercises Fun Activities Games Icebreakers Oneonone Activiti 42747Document2 pagesNatural Disasters Vocabulary Exercises Fun Activities Games Icebreakers Oneonone Activiti 42747Andrea Tercero VillarroelPas encore d'évaluation
- Fpe QB 23Document7 pagesFpe QB 23manishchalke65Pas encore d'évaluation
- Leadership Games and ActivitiesDocument38 pagesLeadership Games and ActivitiesWilliam Oliss100% (1)
- Relationships, 365 Day Devotional Mylesunroe 377pgDocument377 pagesRelationships, 365 Day Devotional Mylesunroe 377pgEla100% (7)
- Intel Stratix 10 Avalon - MM Interface For PCI Express Solutions User GuideDocument173 pagesIntel Stratix 10 Avalon - MM Interface For PCI Express Solutions User Guideenoch richardPas encore d'évaluation
- 1986 EMT Disruption of Microbial Cells For Intracellular ProductsDocument11 pages1986 EMT Disruption of Microbial Cells For Intracellular ProductsLezid CortesPas encore d'évaluation