Vous aimerez peut-être aussi
- Competition & Luxury Vehicle Club of Darlington SuitDocument31 pagesCompetition & Luxury Vehicle Club of Darlington SuitBenjamin DuerPas encore d'évaluation
- Exotic TarantulasDocument82 pagesExotic TarantulasThiago MartinsPas encore d'évaluation
- Report BeetrootDocument11 pagesReport BeetrootSya Subi100% (3)
- Composite Materials: Cris Arnold Materials Research Centre J.c.arnold@swansea - Ac.ukDocument30 pagesComposite Materials: Cris Arnold Materials Research Centre J.c.arnold@swansea - Ac.ukMazin MohammedPas encore d'évaluation
- 몽중1 P&ID 131227-제본파일 (링크 마크업Document272 pages몽중1 P&ID 131227-제본파일 (링크 마크업Lê Thành Chung100% (3)
- Surface Chemistry of Solid and Liquid Interfaces PDFDocument365 pagesSurface Chemistry of Solid and Liquid Interfaces PDFSuchat KotcheapPas encore d'évaluation
- Stimulation Manual ONGC 2008Document306 pagesStimulation Manual ONGC 2008rahul_storm100% (3)
- Steelstrong: Always Strong in Valve WorldDocument32 pagesSteelstrong: Always Strong in Valve WorldBharat Bhushan SharmaPas encore d'évaluation
- Psychedelic LichensDocument4 pagesPsychedelic LichensJon. . .Pas encore d'évaluation
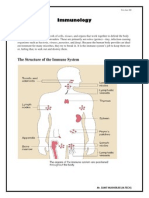
- Immunology NotesDocument8 pagesImmunology NotesSumit Mukherjee100% (1)
- Compannot 101616Document8 pagesCompannot 101616api-320970548Pas encore d'évaluation
- The Life Histories of GenesDocument3 pagesThe Life Histories of Genesjyotirmoy21Pas encore d'évaluation
- 1002 FullDocument14 pages1002 FullcubanosPas encore d'évaluation
- Review: Evolution of Transcriptional Regulatory Circuits in BacteriaDocument12 pagesReview: Evolution of Transcriptional Regulatory Circuits in BacteriajazorelPas encore d'évaluation
- Panchy Et Al (2016) - Plant PhysiologyDocument23 pagesPanchy Et Al (2016) - Plant PhysiologyAna Luiza Atella de FreitasPas encore d'évaluation
- Giriraj Prajapati - Elements of Animal Genetics (2016, Centrum Press) - Libgen - LiDocument225 pagesGiriraj Prajapati - Elements of Animal Genetics (2016, Centrum Press) - Libgen - LiЖеня КоноровPas encore d'évaluation
- Geneticvariation PublishedDocument7 pagesGeneticvariation PublishedTJPlayzPas encore d'évaluation
- MolecularDocument3 pagesMolecularfjafar-jonesPas encore d'évaluation
- Fresard EpigeneticsDocument12 pagesFresard EpigeneticshenkPas encore d'évaluation
- Widespread Whole Genome DuplicationsDocument15 pagesWidespread Whole Genome DuplicationssalarPas encore d'évaluation
- Evolu IntronesDocument5 pagesEvolu IntronesIsaías Turrubiate PPas encore d'évaluation
- Evolutionary Analysis of Endogenous Intronic Retroviruses in Primates Reveals An Enrichment in Transcription Binding Sites Associated With Key Regulatory ProcessesDocument22 pagesEvolutionary Analysis of Endogenous Intronic Retroviruses in Primates Reveals An Enrichment in Transcription Binding Sites Associated With Key Regulatory ProcessesVinura WelgamaPas encore d'évaluation
- Kang Et Al 2014. Adaptative and Nonadaptative Genome SizeDocument11 pagesKang Et Al 2014. Adaptative and Nonadaptative Genome SizelaurachalupPas encore d'évaluation
- 1 s2.0 S1749461320300257 MainDocument8 pages1 s2.0 S1749461320300257 MainBen DresimPas encore d'évaluation
- Thesis EpigeneticsDocument5 pagesThesis Epigeneticsaparnaharrisonstamford100% (2)
- Drosophila: Modeling Tumor Invasion and Metastasis inDocument9 pagesDrosophila: Modeling Tumor Invasion and Metastasis inFernando Lazzarotto EvaldtPas encore d'évaluation
- Running Head: GENETIC DATA 1Document11 pagesRunning Head: GENETIC DATA 1Best EssaysPas encore d'évaluation
- The Prokaryotic Species 2020Document29 pagesThe Prokaryotic Species 2020خديجة بيوPas encore d'évaluation
- Articulo 5Document12 pagesArticulo 5Holman SanabriaPas encore d'évaluation
- Gender Affirming Hormone Therapy Induces Specific DNA Methylation ChangesDocument34 pagesGender Affirming Hormone Therapy Induces Specific DNA Methylation ChangesVallejo Romo Alberto CristianPas encore d'évaluation
- A Multispecies Coalescent Model For Quantitative TraitsDocument24 pagesA Multispecies Coalescent Model For Quantitative Traitshomo sapyensPas encore d'évaluation
- Heritability of Oral Microbial Species in Caries-Active and Caries-FreeDocument13 pagesHeritability of Oral Microbial Species in Caries-Active and Caries-FreeJing XuePas encore d'évaluation
- Elife 68623 v2Document28 pagesElife 68623 v2MarPas encore d'évaluation
- L BosniacumDocument13 pagesL BosniacumStarčević JasminPas encore d'évaluation
- Infection, Genetics and Evolution: Haemaphysalis QinghaiensisDocument15 pagesInfection, Genetics and Evolution: Haemaphysalis QinghaiensisChickencito PerezPas encore d'évaluation
- Research Paper EpigeneticsDocument7 pagesResearch Paper Epigeneticsxcjfderif100% (1)
- Herrera 2011Document14 pagesHerrera 2011Anjar Tri WibowoPas encore d'évaluation
- v1 457252 PDFDocument41 pagesv1 457252 PDFAbhishek YadavPas encore d'évaluation
- Unit 3 (1) ..Document4 pagesUnit 3 (1) ..Valentina RamírezPas encore d'évaluation
- Alu-Mediated Phylogenetic Novelties in Gene Regulation and DevelopmentDocument9 pagesAlu-Mediated Phylogenetic Novelties in Gene Regulation and DevelopmentArtyumR IIPas encore d'évaluation
- SingDocument39 pagesSingThe TruckPas encore d'évaluation
- Fungal DNA BarcodingDocument37 pagesFungal DNA BarcodingAshutosh MukherjeePas encore d'évaluation
- Comparing GenomesDocument11 pagesComparing GenomesAman JainPas encore d'évaluation
- Genetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsDocument13 pagesGenetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsSevim AkçağlarPas encore d'évaluation
- Garcia 2020Document14 pagesGarcia 2020Laura Evangelina GarciaPas encore d'évaluation
- Hormones and Behavior: W. Lee, M.F. Dwortz, T.M. Milewski, F.A. Champagne, J.P. CurleyDocument11 pagesHormones and Behavior: W. Lee, M.F. Dwortz, T.M. Milewski, F.A. Champagne, J.P. CurleyAuxane FaugierPas encore d'évaluation
- Group 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroDocument4 pagesGroup 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroAnne Olfato ParojinogPas encore d'évaluation
- Evolutionary Mechanisms of Microbial GenomesDocument131 pagesEvolutionary Mechanisms of Microbial GenomeszrpyhjtzztPas encore d'évaluation
- Apprehending Multicellularity: Regulatory Networks, Genomics, and EvolutionDocument22 pagesApprehending Multicellularity: Regulatory Networks, Genomics, and EvolutionsomasushmaPas encore d'évaluation
- BMC Evolutionary Biology: Unique Genes in Plants: Specificities and Conserved Features Throughout EvolutionDocument20 pagesBMC Evolutionary Biology: Unique Genes in Plants: Specificities and Conserved Features Throughout EvolutionRachanaPas encore d'évaluation
- Gene Polymorphisms and Male Infertility - A Meta-Analysis and Literature ReviewDocument8 pagesGene Polymorphisms and Male Infertility - A Meta-Analysis and Literature ReviewafmzdabdjteegfPas encore d'évaluation
- Developmental Biology: Abigail M. Anderson, Brandon P. Weasner, Bonnie M. Weasner, Justin P. KumarDocument11 pagesDevelopmental Biology: Abigail M. Anderson, Brandon P. Weasner, Bonnie M. Weasner, Justin P. KumartinaPas encore d'évaluation
- Research Paper On Plant BiotechnologyDocument7 pagesResearch Paper On Plant Biotechnologyafnkaufhczyvbc100% (1)
- Budddevosfmicb 03 00401Document10 pagesBudddevosfmicb 03 00401Aidan BuddPas encore d'évaluation
- STRA65Document6 pagesSTRA65SUJITH232323Pas encore d'évaluation
- Comp Transcript Between Zebrafsh and MammalsDocument5 pagesComp Transcript Between Zebrafsh and MammalsRita RodriguesPas encore d'évaluation
- The Relationship Between The Gut Microbiome and Host Gene Expression: A ReviewDocument14 pagesThe Relationship Between The Gut Microbiome and Host Gene Expression: A ReviewsznistvanPas encore d'évaluation
- Comparative Biochemistry and Physiology Part A: Molecular & Integrative PhysiologyDocument9 pagesComparative Biochemistry and Physiology Part A: Molecular & Integrative PhysiologyАлександр КолядаPas encore d'évaluation
- TMP F780Document9 pagesTMP F780FrontiersPas encore d'évaluation
- Developmental Bias in Horned Dung Beetles and Its Contributions To Innovation, Adaptation, and ResilienceDocument16 pagesDevelopmental Bias in Horned Dung Beetles and Its Contributions To Innovation, Adaptation, and ResilienceelodrasPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument9 pagesNIH Public Access: Author ManuscriptAndressa AlvesPas encore d'évaluation
- Dimorfismo SexualDocument21 pagesDimorfismo SexualDiana CortesPas encore d'évaluation
- Tmpa688 TMPDocument12 pagesTmpa688 TMPFrontiersPas encore d'évaluation
- The Generation of Animal Diversity: Word Count: 1088Document6 pagesThe Generation of Animal Diversity: Word Count: 1088api-345977843Pas encore d'évaluation
- Trabajo InglesDocument3 pagesTrabajo InglesValentina RamírezPas encore d'évaluation
- GenqDocument10 pagesGenqTankguh RevolusionerPas encore d'évaluation
- Evolution of Human Diseases - HO Smail PDFDocument17 pagesEvolution of Human Diseases - HO Smail PDFdoeditPas encore d'évaluation
- Homossexualidade e GeneticaDocument27 pagesHomossexualidade e GeneticaMônica AlvesPas encore d'évaluation
- Mutations in Human Accelerated Regions Disrupt Cognition and Social BehaviorDocument27 pagesMutations in Human Accelerated Regions Disrupt Cognition and Social BehaviorJiayao HuangPas encore d'évaluation
- Clinical Genetics and Genomics of AgingD'EverandClinical Genetics and Genomics of AgingJuan Carlos Gomez-VerjanPas encore d'évaluation
- The Hepatitis B Virus Pres1 Domain Hijacks Host Trafficking Proteins by Motif MimicryDocument10 pagesThe Hepatitis B Virus Pres1 Domain Hijacks Host Trafficking Proteins by Motif MimicrySumit MukherjeePas encore d'évaluation
- Effector ReviewDocument26 pagesEffector ReviewSumit MukherjeePas encore d'évaluation
- A Tale of Effectors Their Secretory Mechanisms and Computational Discovery 2168 9547-3-118Document14 pagesA Tale of Effectors Their Secretory Mechanisms and Computational Discovery 2168 9547-3-118Sumit MukherjeePas encore d'évaluation
- NR Micro 1549Document10 pagesNR Micro 1549Sumit MukherjeePas encore d'évaluation
- Bio-Liinux Nature PDFDocument3 pagesBio-Liinux Nature PDFSumit MukherjeePas encore d'évaluation
- Notification NICL Administrative OfficersDocument7 pagesNotification NICL Administrative OfficersRushi BagePas encore d'évaluation
- N R SuggestionDocument2 pagesN R SuggestionSumit MukherjeePas encore d'évaluation
- Clinical Restorative Dental Materials Guide: University of California Los Angeles School of DentistryDocument43 pagesClinical Restorative Dental Materials Guide: University of California Los Angeles School of DentistrySergioPachecoSerranoPas encore d'évaluation
- AnchorDocument5 pagesAnchorpadalakirankumarPas encore d'évaluation
- Nabl Accreditation LetterDocument2 pagesNabl Accreditation LetterAnubhav LakhmaniPas encore d'évaluation
- Langbeinite ProcessingDocument5 pagesLangbeinite ProcessingPınar EroğluPas encore d'évaluation
- Properties of WaterDocument18 pagesProperties of WaterJay Vincent MacadangdangPas encore d'évaluation
- Analisis Fitokimia Daun Pepaya (Carica Papaya L.) A'yun Et Al.Document7 pagesAnalisis Fitokimia Daun Pepaya (Carica Papaya L.) A'yun Et Al.Cahaya Medika1Pas encore d'évaluation
- Research Proposal - Chaper 1Document10 pagesResearch Proposal - Chaper 1Kia Javar100% (1)
- SF6 Sulfur Hexafluoride PDFDocument12 pagesSF6 Sulfur Hexafluoride PDFSujit Kumar pahiPas encore d'évaluation
- Intro AzoDyeDocument2 pagesIntro AzoDyejammPas encore d'évaluation
- Edwards CP25K Cold Cathode Gauge Sensor ManualDocument18 pagesEdwards CP25K Cold Cathode Gauge Sensor Manualঅর্ণব কোলেPas encore d'évaluation
- Cambridge International AS & A Level: Chemistry 9701/12Document16 pagesCambridge International AS & A Level: Chemistry 9701/12O and A Level TutorPas encore d'évaluation
- Emerging Solar Cell PDFDocument23 pagesEmerging Solar Cell PDFJaffar LonePas encore d'évaluation
- Simplified Construction Estimate: ConcreteDocument11 pagesSimplified Construction Estimate: ConcreteGreta Andrea TalipPas encore d'évaluation
- Sika Injection 20Document3 pagesSika Injection 20the pilotPas encore d'évaluation
- Iso AnnealingDocument2 pagesIso AnnealingPurushottam Sutar100% (1)
- BeerDocument21 pagesBeermichol20140% (1)
- Principles of Food Processing and Halal Requirement (WEEK 11)Document48 pagesPrinciples of Food Processing and Halal Requirement (WEEK 11)aishah1997Pas encore d'évaluation
- Buku Miller's Anesthesia BAB 20 Dan 21Document88 pagesBuku Miller's Anesthesia BAB 20 Dan 21Rahma TomuPas encore d'évaluation
- Module 1Document15 pagesModule 1venugopal_aeroPas encore d'évaluation
- Principles of Soil Science Exercise Manual: Mark W. BowenDocument70 pagesPrinciples of Soil Science Exercise Manual: Mark W. BowenDominic ImperialePas encore d'évaluation
- Group #4: September 25, 2019Document5 pagesGroup #4: September 25, 2019MIKAYLA ELAINE P. DELA CRUZPas encore d'évaluation