Vous aimerez peut-être aussi
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- 01 Moore Eval BuccalDocument7 pages01 Moore Eval BuccalRoy WilsonPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- ArticleDocument8 pagesArticleRoy WilsonPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Peroxisomal β-oxidation-A metabolic pathway with multiple functionsDocument14 pagesPeroxisomal β-oxidation-A metabolic pathway with multiple functionsRoy WilsonPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- J Med Genet 2004 Khan E79 - 2Document7 pagesJ Med Genet 2004 Khan E79 - 2Roy WilsonPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Peroxisome Proliferator Activated Receptors: Transcriptional Regulators of Adipogenesis, Lipid Metabolism and More..Document6 pagesPeroxisome Proliferator Activated Receptors: Transcriptional Regulators of Adipogenesis, Lipid Metabolism and More..Roy WilsonPas encore d'évaluation
- 176 332 1 SMDocument10 pages176 332 1 SMRoy WilsonPas encore d'évaluation
- Fahey Lab 04 02Document4 pagesFahey Lab 04 02Roy WilsonPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Hennis Et Al Genetic Factors Associated With Exercise Performance in Atmospheric Hypoxia 2015 Online VersionDocument17 pagesHennis Et Al Genetic Factors Associated With Exercise Performance in Atmospheric Hypoxia 2015 Online VersionRoy WilsonPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Strength Training For Endurance Athletes Theory.1Document12 pagesStrength Training For Endurance Athletes Theory.1Roy WilsonPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Short-Term Power Output and Local Muscular Endurance of Young Male Soccer Players According To Playing PositionDocument7 pagesShort-Term Power Output and Local Muscular Endurance of Young Male Soccer Players According To Playing PositionRoy WilsonPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- C34T Mutation of The AMPD1 Gene in An Elite White Runner: Case ReportDocument3 pagesC34T Mutation of The AMPD1 Gene in An Elite White Runner: Case ReportRoy WilsonPas encore d'évaluation
- Myoadenylate Deaminase Deficiency Associated To A Liver Cirrhosis: A Possible Cause of Neurological Dysfunction Resembling A Hepatic EncephalopathyDocument4 pagesMyoadenylate Deaminase Deficiency Associated To A Liver Cirrhosis: A Possible Cause of Neurological Dysfunction Resembling A Hepatic EncephalopathyRoy WilsonPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- 1 Hadzovic DzuvoDocument7 pages1 Hadzovic DzuvoRoy WilsonPas encore d'évaluation
- Intense and Exhaustive Exercise Induce Oxidative Stress in Skeletal MuscleDocument4 pagesIntense and Exhaustive Exercise Induce Oxidative Stress in Skeletal MuscleRoy WilsonPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Mitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingDocument7 pagesMitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingRoy WilsonPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Lipid Peroxidation Inhibition and Antiradical Activities of Some Leaf Fractions of Mangifera IndicaDocument7 pagesLipid Peroxidation Inhibition and Antiradical Activities of Some Leaf Fractions of Mangifera IndicaRoy WilsonPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Comparison of The Effects of A Single Bout of Resistance Exercises With Different Intensities On Oxidative Stress in Untrained Male StudentsDocument5 pagesComparison of The Effects of A Single Bout of Resistance Exercises With Different Intensities On Oxidative Stress in Untrained Male StudentsRoy WilsonPas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Tackling Food Inflation: Ashwinkumar Kokku - 67 Malcolm Pinto - 89 Samir Vele - Nitin JadhavDocument9 pagesTackling Food Inflation: Ashwinkumar Kokku - 67 Malcolm Pinto - 89 Samir Vele - Nitin JadhavMalcolm PintoPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Distress Manual PDFDocument51 pagesDistress Manual PDFEIRINI ZIGKIRIADOUPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Hamraki Rag April 2010 IssueDocument20 pagesHamraki Rag April 2010 IssueHamraki RagPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Case Report 3 MukokelDocument3 pagesCase Report 3 MukokelWidychii GadiestchhetyaPas encore d'évaluation
- E-Kabin - O Series - Monoblock Enclosure - ENGDocument12 pagesE-Kabin - O Series - Monoblock Enclosure - ENGCatalina CocoşPas encore d'évaluation
- Family Stress TheoryDocument10 pagesFamily Stress TheoryKarina Megasari WinahyuPas encore d'évaluation
- L A R G e - S C A L e M e T H A M P H e T A M I N e M A N U F A C T U R eDocument21 pagesL A R G e - S C A L e M e T H A M P H e T A M I N e M A N U F A C T U R eDaria Schka100% (1)
- Catalogue CV. Traka Abadi UniversalDocument15 pagesCatalogue CV. Traka Abadi UniversalHackers StevenPas encore d'évaluation
- Doingwell Coaching ServicesDocument4 pagesDoingwell Coaching ServicesPaweł PawełPas encore d'évaluation
- Amul Amul AmulDocument7 pagesAmul Amul Amulravikumarverma28Pas encore d'évaluation
- Intentions and Results ASFA and Incarcerated ParentsDocument10 pagesIntentions and Results ASFA and Incarcerated Parentsaflee123Pas encore d'évaluation
- Overall Summary:: SAP MM Certified Associate & SAP Certification ID: 0019350978Document6 pagesOverall Summary:: SAP MM Certified Associate & SAP Certification ID: 0019350978Ganapathi RajPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- ANATOMY 1 NILEM and Cat FISHDocument19 pagesANATOMY 1 NILEM and Cat FISHAnisatul Khabibah ZaenPas encore d'évaluation
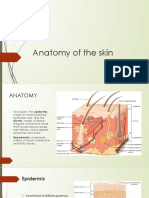
- Anatomy of The SkinDocument28 pagesAnatomy of The Skinay254Pas encore d'évaluation
- Đề ANH chuyên 5Document7 pagesĐề ANH chuyên 5Phạm Ngô Hiền MaiPas encore d'évaluation
- Hemostatic AgentsDocument18 pagesHemostatic AgentshariPas encore d'évaluation
- Schedule NDocument3 pagesSchedule Nmittal kelaPas encore d'évaluation
- Essay 31 - Permissive ParentingDocument2 pagesEssay 31 - Permissive Parentingqbich37Pas encore d'évaluation
- Ancon Tension Systems March 2008Document16 pagesAncon Tension Systems March 2008Slinky BillPas encore d'évaluation
- UgpeDocument3 pagesUgpeOlety Subrahmanya SastryPas encore d'évaluation
- 4 StartUp GuideDocument2 pages4 StartUp GuideSamuel RomeroPas encore d'évaluation
- A Cook's Journey To Japan - Fish Tales and Rice Paddies 100 Homestyle Recipes From Japanese KitchensDocument306 pagesA Cook's Journey To Japan - Fish Tales and Rice Paddies 100 Homestyle Recipes From Japanese KitchensEthan F.100% (1)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Universal ING - LA.Boschi Plants Private LimitedDocument23 pagesUniversal ING - LA.Boschi Plants Private LimitedAlvaro Mendoza MaytaPas encore d'évaluation
- Experiment Number 16 Formal ReportDocument4 pagesExperiment Number 16 Formal Reportapi-524547303Pas encore d'évaluation
- Astm d2729Document2 pagesAstm d2729Shan AdriasPas encore d'évaluation
- Aakanksha ProjectDocument32 pagesAakanksha ProjectAakash murarkaPas encore d'évaluation
- Carolyn Green Release FinalDocument3 pagesCarolyn Green Release FinalAlex MilesPas encore d'évaluation
- Remote Control Unit Manual BookDocument21 pagesRemote Control Unit Manual BookIgor Ungur100% (1)
- 2021 Life Skills ATP Grade 5Document10 pages2021 Life Skills ATP Grade 5Koketso SekwenyanePas encore d'évaluation
- Manual of GardeningDocument812 pagesManual of GardeningPrakash PatelPas encore d'évaluation
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceD'EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceÉvaluation : 4.5 sur 5 étoiles4.5/5 (517)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseD'EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseÉvaluation : 4.5 sur 5 étoiles4.5/5 (52)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4 sur 5 étoiles4/5 (5)