Vous aimerez peut-être aussi
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsD'EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochranePas encore d'évaluation
- The Molecular Mechanisms of Glucocorticoids-Mediated Neutrophil SurvivalDocument7 pagesThe Molecular Mechanisms of Glucocorticoids-Mediated Neutrophil Survivalade_liaPas encore d'évaluation
- Basic Biology and Clinical Aspects of InflammationD'EverandBasic Biology and Clinical Aspects of InflammationPas encore d'évaluation
- NETosis: Molecular Mechanisms, Role in Physiology and PathologyDocument22 pagesNETosis: Molecular Mechanisms, Role in Physiology and PathologysebasgaravanoPas encore d'évaluation
- Inflammation: From Molecular and Cellular Mechanisms to the ClinicD'EverandInflammation: From Molecular and Cellular Mechanisms to the ClinicJean-Marc CavaillonPas encore d'évaluation
- Ijcep0011 4283Document13 pagesIjcep0011 4283tomalbertoelgoldenPas encore d'évaluation
- Bayazid 2021Document11 pagesBayazid 2021Jocilene Dantas Torres NascimentoPas encore d'évaluation
- Neutrophil in Chronic InflammationDocument15 pagesNeutrophil in Chronic InflammationRizqaFebrilianyPas encore d'évaluation
- 1 s2.0 S0021925820870148 MainDocument11 pages1 s2.0 S0021925820870148 MainAina SarahPas encore d'évaluation
- 2007, Vol.25, Issues 4, Cutaneous Receptors - Clinical Implications and Therapeutic RelevanceDocument202 pages2007, Vol.25, Issues 4, Cutaneous Receptors - Clinical Implications and Therapeutic RelevanceRizweta DestinPas encore d'évaluation
- Gupta2014 - NETs Calcium CyclosporineA - PLoS OneDocument12 pagesGupta2014 - NETs Calcium CyclosporineA - PLoS OneHans MenaresPas encore d'évaluation
- Necroptosis PDFDocument11 pagesNecroptosis PDFdavid zambranoPas encore d'évaluation
- 1 s2.0 S0085253815474071 MainDocument8 pages1 s2.0 S0085253815474071 MainBintiPas encore d'évaluation
- RIP Kinases: Key Decision Makers in Cell Death and Innate ImmunityDocument12 pagesRIP Kinases: Key Decision Makers in Cell Death and Innate ImmunityBalarama Krishna GogineniPas encore d'évaluation
- Nat Rev Immunol 2011 WestDocument15 pagesNat Rev Immunol 2011 WestGabriel CamarenaPas encore d'évaluation
- Belas Figuras Nada de CampyDocument6 pagesBelas Figuras Nada de CampyHaurusPas encore d'évaluation
- Mtb TLR TNFα NFkBDocument9 pagesMtb TLR TNFα NFkBamwayindia2024Pas encore d'évaluation
- Sepsis y CID 2016Document8 pagesSepsis y CID 2016Ellys Macías PeraltaPas encore d'évaluation
- Nets X Fungos 2019Document24 pagesNets X Fungos 2019Bruno Nogueira BarrosPas encore d'évaluation
- Tnf-A Protein Synthesis Inhibitor Restores Neuronal Function and Reverses Cognitive Deficits Induced by Chronic NeuroinflammationDocument13 pagesTnf-A Protein Synthesis Inhibitor Restores Neuronal Function and Reverses Cognitive Deficits Induced by Chronic NeuroinflammationAlex AlexPas encore d'évaluation
- Toll-Like Receptors: The Swiss Army Knife of Immunity and Vaccine DevelopmentDocument10 pagesToll-Like Receptors: The Swiss Army Knife of Immunity and Vaccine DevelopmentWilson Barros LuizPas encore d'évaluation
- Quinderé Et Al 2015 PDFDocument9 pagesQuinderé Et Al 2015 PDFLuizaPas encore d'évaluation
- Role of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsDocument10 pagesRole of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsAsish GeiorgePas encore d'évaluation
- Bacterial Meningitis 19Document10 pagesBacterial Meningitis 19caliptra36Pas encore d'évaluation
- Cells 10 02406Document18 pagesCells 10 02406Tung NguyenPas encore d'évaluation
- Infeccion MicrobianaDocument18 pagesInfeccion MicrobianaRosemary Neto AlfonsoPas encore d'évaluation
- Reviews: Neutrophil Diversity and Plasticity in Tumour Progression and TherapyDocument19 pagesReviews: Neutrophil Diversity and Plasticity in Tumour Progression and TherapySobek1789Pas encore d'évaluation
- Neutrophil Extracellular Traps Exert Potential Cytotoxic and Proinflammatory Effects in The Dental PulpDocument11 pagesNeutrophil Extracellular Traps Exert Potential Cytotoxic and Proinflammatory Effects in The Dental PulpPentiya Vita AyuniPas encore d'évaluation
- Apoptotic Clearance RegulationDocument15 pagesApoptotic Clearance Regulationare4erPas encore d'évaluation
- The Pseudokinase MLKL Activates PAD4-dependent NETDocument27 pagesThe Pseudokinase MLKL Activates PAD4-dependent NETshaheena ahmedPas encore d'évaluation
- Carestia 2015Document10 pagesCarestia 2015Ricardo GomezPas encore d'évaluation
- tmp4C1C TMPDocument8 pagestmp4C1C TMPFrontiersPas encore d'évaluation
- tmpF65B TMPDocument8 pagestmpF65B TMPFrontiersPas encore d'évaluation
- Modelamiento Matemàtico de La ApoptosisDocument7 pagesModelamiento Matemàtico de La ApoptosisAlexisPas encore d'évaluation
- Net InductionDocument18 pagesNet InductionManovriti ThakurPas encore d'évaluation
- Pathophysiologyofsepsis Up To DateDocument24 pagesPathophysiologyofsepsis Up To DateTom CoutinhoPas encore d'évaluation
- PatogenesisDocument4 pagesPatogenesisFajar Cristianta GintingPas encore d'évaluation
- 1 Melatonin Prevents Allergic Airway Inflammation in Epicutaneously Sensitized MiceDocument12 pages1 Melatonin Prevents Allergic Airway Inflammation in Epicutaneously Sensitized Micebemfisio.cristianeribeiroPas encore d'évaluation
- Neutrophils at The Crossroads of Innate and Adaptive Immunity. J Leukoc Biol. 2020 JulDocument20 pagesNeutrophils at The Crossroads of Innate and Adaptive Immunity. J Leukoc Biol. 2020 Julantonychino10Pas encore d'évaluation
- NF KB DissertationDocument5 pagesNF KB DissertationBuyPapersForCollegeAlbuquerque100% (1)
- Yu 2012Document9 pagesYu 2012KianaPas encore d'évaluation
- Anti Inflammatory PhytochemicalsDocument14 pagesAnti Inflammatory PhytochemicalsMichael Kohlberger, BSc MScPas encore d'évaluation
- Protein Kinase C-Delta Regulates HIV-1 Replication at An Early Post-Entry Step in MacrophagesDocument11 pagesProtein Kinase C-Delta Regulates HIV-1 Replication at An Early Post-Entry Step in MacrophagesMarliCurcioPas encore d'évaluation
- Immunologyoffungal Infections: Oscar A. Fernández-García,, Jennifer M. Cuellar-RodríguezDocument16 pagesImmunologyoffungal Infections: Oscar A. Fernández-García,, Jennifer M. Cuellar-RodríguezsilviaPas encore d'évaluation
- Neutrophils Induce Paracrine Telomere Dysfunction and Senescence in ROS-dependent MannerDocument19 pagesNeutrophils Induce Paracrine Telomere Dysfunction and Senescence in ROS-dependent MannerValen EstevezPas encore d'évaluation
- Macrophage Response Towards LPSDocument10 pagesMacrophage Response Towards LPSajaewoongPas encore d'évaluation
- Diagnostics 12 02004 v2Document12 pagesDiagnostics 12 02004 v2Appiah Peter OforiPas encore d'évaluation
- Inmunología 4Document8 pagesInmunología 4Valentina PereiraPas encore d'évaluation
- Pathogenesis of Leptospirosis Important Issues 2161 0703.1000e127 1Document2 pagesPathogenesis of Leptospirosis Important Issues 2161 0703.1000e127 1Anonymous 7jKR9XbPas encore d'évaluation
- 1 s2.0 S0165247822000645 MainDocument7 pages1 s2.0 S0165247822000645 Mainnatacatt13Pas encore d'évaluation
- Patho Case 1Document12 pagesPatho Case 1Venz Timothy Wesley LandichoPas encore d'évaluation
- 2019 Article 302Document11 pages2019 Article 302Noy FaridaPas encore d'évaluation
- Cytokines and DiseaseDocument2 pagesCytokines and DiseaseGeorgiGugicevPas encore d'évaluation
- Neutrophils in Periodontal DiseaseDocument35 pagesNeutrophils in Periodontal DiseaseVishali RayapudiPas encore d'évaluation
- Fimmu 11 585880Document14 pagesFimmu 11 585880Irvin MarcelPas encore d'évaluation
- Thalidomide and Celecoxib For Severe COVID-19 PneumoniaDocument11 pagesThalidomide and Celecoxib For Severe COVID-19 Pneumonialptscsi100% (1)
- Etiology InflammationDocument15 pagesEtiology InflammationYeni RahmawatiPas encore d'évaluation
- Targeting Neddylation Pathway With MLN4924 (Pevonedistat) InducesDocument6 pagesTargeting Neddylation Pathway With MLN4924 (Pevonedistat) InducescarlakerengrPas encore d'évaluation
- Nri 1936 ArtigoDocument13 pagesNri 1936 ArtigoGlauce L TrevisanPas encore d'évaluation
- An 1Document10 pagesAn 1Faqih SultonfaqPas encore d'évaluation
- Intestinal Nematodes (Unholy Trinity)Document6 pagesIntestinal Nematodes (Unholy Trinity)DAN JR. M. BAUSAPas encore d'évaluation
- Anaphy Lec Sas-17Document4 pagesAnaphy Lec Sas-17Francis Steve RipdosPas encore d'évaluation
- Homoeopathic Management of Chronic SinusitisDocument6 pagesHomoeopathic Management of Chronic SinusitisEditor IJTSRDPas encore d'évaluation
- Infection MCQDocument49 pagesInfection MCQalh basharPas encore d'évaluation
- Parasitology Guide QuestionDocument2 pagesParasitology Guide QuestionRoll BiPas encore d'évaluation
- Allergen Calendar - EnglishDocument2 pagesAllergen Calendar - Englishpkgill15Pas encore d'évaluation
- 5.0 LeukopoiesisDocument37 pages5.0 LeukopoiesisJunior SataPas encore d'évaluation
- Vaccination CentersDocument4 pagesVaccination CentersdmonlinelkPas encore d'évaluation
- Burkitt Lymphoma: Teacher: Daniel Hernández. Student: Diana Rosales DominguezDocument16 pagesBurkitt Lymphoma: Teacher: Daniel Hernández. Student: Diana Rosales DominguezDianita Rosales Dominguez100% (1)
- Hemolytic Uremic SyndromeDocument3 pagesHemolytic Uremic SyndromeEliDavid100% (1)
- Elecsys Hbsag Ii: A) Tris (2,2'-Bipyridyl) Ruthenium (Ii) - Complex (Ru (Bpy) )Document5 pagesElecsys Hbsag Ii: A) Tris (2,2'-Bipyridyl) Ruthenium (Ii) - Complex (Ru (Bpy) )Brian SamanyaPas encore d'évaluation
- NICU Manual Cheat SheetDocument2 pagesNICU Manual Cheat Sheetlori_quintal92% (13)
- Food Allergy TrainingDocument10 pagesFood Allergy TrainingHakim AliPas encore d'évaluation
- Dr. Reviono, DR., SP.P (K) - Update Diagnosis and Management JeroveciDocument20 pagesDr. Reviono, DR., SP.P (K) - Update Diagnosis and Management JeroveciOlivia DwimaswastiPas encore d'évaluation
- EUCAST Breakpoints V 2.0 120101Document73 pagesEUCAST Breakpoints V 2.0 120101mermarinarPas encore d'évaluation
- St. Paul College of Ilocos Sur: Stages in The Natural History of Disease and The Levels of Prevention During Ncov-19Document3 pagesSt. Paul College of Ilocos Sur: Stages in The Natural History of Disease and The Levels of Prevention During Ncov-19Marie Kelsey Acena Macaraig100% (1)
- EncephalitisDocument30 pagesEncephalitisHITIPHYSIO100% (1)
- Sure Vue ASO Pi PDFDocument4 pagesSure Vue ASO Pi PDFArifani CahyaniPas encore d'évaluation
- Extrapulmonary TuberculosisDocument80 pagesExtrapulmonary TuberculosisYoursTruly FIBONPas encore d'évaluation
- Pathophysiology ImpetigoDocument3 pagesPathophysiology ImpetigoAnne de VeraPas encore d'évaluation
- Introduction To ImmunologyDocument10 pagesIntroduction To ImmunologyArvi MandawePas encore d'évaluation
- EUA Roche Elecsys IfuDocument5 pagesEUA Roche Elecsys IfuFadhila N HandayaniPas encore d'évaluation
- Acute Pancreatitis PresentationDocument24 pagesAcute Pancreatitis PresentationhtmgPas encore d'évaluation
- Oral Candidiasis: Madhu Priya.MDocument4 pagesOral Candidiasis: Madhu Priya.MinventionjournalsPas encore d'évaluation
- Bombay Blood GroupDocument1 pageBombay Blood GroupSanjay GuptaPas encore d'évaluation
- Epidemiology Classes PDFDocument167 pagesEpidemiology Classes PDFsathish kannamPas encore d'évaluation
- B.B Immunology For Blood BankDocument26 pagesB.B Immunology For Blood BankAbode AlharbiPas encore d'évaluation
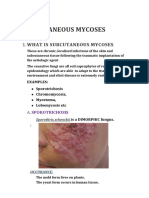
- What Is Subcutaneous MycosesDocument4 pagesWhat Is Subcutaneous MycosesArslan SaeedPas encore d'évaluation
- PDF TextDocument2 pagesPDF TextSATYAM KANNAUJPas encore d'évaluation
- Vaccine Rejection and HesitancyDocument7 pagesVaccine Rejection and HesitancyAlexander FrischPas encore d'évaluation
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionD'EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionÉvaluation : 4 sur 5 étoiles4/5 (404)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDD'EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDÉvaluation : 5 sur 5 étoiles5/5 (3)
- The Age of Magical Overthinking: Notes on Modern IrrationalityD'EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityÉvaluation : 4 sur 5 étoiles4/5 (29)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeD'EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeÉvaluation : 2 sur 5 étoiles2/5 (1)
- The Obesity Code: Unlocking the Secrets of Weight LossD'EverandThe Obesity Code: Unlocking the Secrets of Weight LossÉvaluation : 4 sur 5 étoiles4/5 (6)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)D'EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Évaluation : 3 sur 5 étoiles3/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisD'EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (42)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedD'EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedÉvaluation : 5 sur 5 étoiles5/5 (81)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsD'EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsPas encore d'évaluation
- The Comfort of Crows: A Backyard YearD'EverandThe Comfort of Crows: A Backyard YearÉvaluation : 4.5 sur 5 étoiles4.5/5 (23)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.D'EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Évaluation : 4.5 sur 5 étoiles4.5/5 (110)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsD'EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessD'EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessÉvaluation : 4.5 sur 5 étoiles4.5/5 (328)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaD'EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- To Explain the World: The Discovery of Modern ScienceD'EverandTo Explain the World: The Discovery of Modern ScienceÉvaluation : 3.5 sur 5 étoiles3.5/5 (51)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4 sur 5 étoiles4/5 (5)
- Troubled: A Memoir of Foster Care, Family, and Social ClassD'EverandTroubled: A Memoir of Foster Care, Family, and Social ClassÉvaluation : 4.5 sur 5 étoiles4.5/5 (27)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeD'EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeÉvaluation : 4.5 sur 5 étoiles4.5/5 (253)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisD'EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisÉvaluation : 4 sur 5 étoiles4/5 (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsD'EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsÉvaluation : 4 sur 5 étoiles4/5 (4)
- The Marshmallow Test: Mastering Self-ControlD'EverandThe Marshmallow Test: Mastering Self-ControlÉvaluation : 4.5 sur 5 étoiles4.5/5 (59)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsD'EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsÉvaluation : 4.5 sur 5 étoiles4.5/5 (170)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryD'EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryÉvaluation : 4 sur 5 étoiles4/5 (45)