Vous aimerez peut-être aussi
- Sistema UrinarioDocument8 pagesSistema UrinarioAlexis S. RuizPas encore d'évaluation
- 7° Prueba Sistema InmuneDocument5 pages7° Prueba Sistema InmuneJimena Velásquez58% (24)
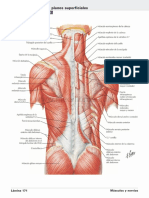
- Musculos de DorsoDocument2 pagesMusculos de Dorsojhon jairo tarapues cuaycal100% (1)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017D'EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Pas encore d'évaluation
- Clase de Biología para PAES.Document19 pagesClase de Biología para PAES.Belen antonia torresPas encore d'évaluation
- Lecciones de inmunología esencialesDocument32 pagesLecciones de inmunología esencialesDaniel Mendoza100% (1)
- Examen Fisico PreocupacionalDocument4 pagesExamen Fisico Preocupacionalclinica ato rtbPas encore d'évaluation
- Las Implicaciones de La Hipermutacion SomaticaDocument6 pagesLas Implicaciones de La Hipermutacion SomaticaGianella PachecoPas encore d'évaluation
- Preguntas para La Unidad 3 de Inmunología Referente A Órganos y Células LinfoidesDocument4 pagesPreguntas para La Unidad 3 de Inmunología Referente A Órganos y Células LinfoidesSerCL1Pas encore d'évaluation
- Activación Del B y Producción de AnticuerposDocument3 pagesActivación Del B y Producción de AnticuerposOctavio ZmPas encore d'évaluation
- Activación de Linfocitos T AlorreactivosDocument9 pagesActivación de Linfocitos T AlorreactivosManuel Rojas GonzaalesPas encore d'évaluation
- Inmunología B y respuesta humoralDocument8 pagesInmunología B y respuesta humoralDani RodriguezPas encore d'évaluation
- Determinante Antigénico IDocument6 pagesDeterminante Antigénico IKaren MendiolaPas encore d'évaluation
- Inmuno 2do ParcialDocument3 pagesInmuno 2do ParcialLinda MagdalenoPas encore d'évaluation
- OntogeniaDocument19 pagesOntogeniaRocío StürtzPas encore d'évaluation
- Objetivos Capitulo 10 Inmunologia UprrpDocument14 pagesObjetivos Capitulo 10 Inmunologia UprrpAlexandra Leonor Pujols AstacioPas encore d'évaluation
- Resumen Cap 6 InmunoDocument6 pagesResumen Cap 6 Inmunobopeepout papuPas encore d'évaluation
- Respuesta Celular y HumoralDocument33 pagesRespuesta Celular y HumoralHugoRafaelGarciaMirandaPas encore d'évaluation
- Activación Del Linfocito B y Producción de AnticuerposDocument6 pagesActivación Del Linfocito B y Producción de AnticuerposJoseMariaMtzPas encore d'évaluation
- Clase 8 Inmunidad Mediada Por CélulasDocument42 pagesClase 8 Inmunidad Mediada Por CélulasYOCELINPas encore d'évaluation
- Prt12 Inm ProlCelDocument12 pagesPrt12 Inm ProlCelGerardo SotoPas encore d'évaluation
- Guia de Estudio de InmunologiaDocument7 pagesGuia de Estudio de InmunologiaAlexei AshfordPas encore d'évaluation
- Taller de Inmuno 23Document4 pagesTaller de Inmuno 23Eliza Moreno SalamancaPas encore d'évaluation
- Investiga RDocument5 pagesInvestiga RCarlos Eduardo Díaz MoralesPas encore d'évaluation
- LLA-B Parte 2: Ontogenia de Linfocitos T y BDocument14 pagesLLA-B Parte 2: Ontogenia de Linfocitos T y BEri MñzPas encore d'évaluation
- Clase 5Document4 pagesClase 5Jesus Yepes martinezPas encore d'évaluation
- Linfocitos T CD4+Document12 pagesLinfocitos T CD4+pitu anatoPas encore d'évaluation
- TOLERANCIA DE LINFOCITOS T Y BDocument6 pagesTOLERANCIA DE LINFOCITOS T Y BKenia espinosa mayerliiiPas encore d'évaluation
- Clase 19.10 Inmunidad Humoral: Funciones de los Ac y Activación de LBDocument14 pagesClase 19.10 Inmunidad Humoral: Funciones de los Ac y Activación de LBElías Damián Martínez TerénPas encore d'évaluation
- Carpeta 5Document7 pagesCarpeta 5katherine MontásPas encore d'évaluation
- Linfocitos TDocument15 pagesLinfocitos TIGNACIO SALVADOR FERNANDEZ GIRONPas encore d'évaluation
- Bloque III 4Document4 pagesBloque III 4Qbp RdrgzPas encore d'évaluation
- Tolerancia y AutoinmunidadDocument8 pagesTolerancia y AutoinmunidadLuis Alberto de la Fuente García PeñaPas encore d'évaluation
- resumen-cap-6-inmunologia-de-abbasDocument7 pagesresumen-cap-6-inmunologia-de-abbasmontsesitacm1104Pas encore d'évaluation
- Inmunología Del TrasplanteDocument8 pagesInmunología Del TrasplanteSOL ESTEPHANIE MELENDEZ GURROLAPas encore d'évaluation
- Linfocitos BDocument5 pagesLinfocitos BLucio WekslerPas encore d'évaluation
- TrasplantesDocument43 pagesTrasplantesSofi Luna 赤い月Pas encore d'évaluation
- Linfocitos 2017Document88 pagesLinfocitos 2017Monica OrdinolaPas encore d'évaluation
- Transplante RenalDocument38 pagesTransplante RenalMELISSA MAGALY SUCASAIRE CASTROPas encore d'évaluation
- RESPUESTA INMUNE CELULAR CompletaDocument96 pagesRESPUESTA INMUNE CELULAR CompletaJazmin Sanchez SanchezPas encore d'évaluation
- Maduración y Activación de Linfocitos TDocument59 pagesMaduración y Activación de Linfocitos TVioleta Tello MedinaPas encore d'évaluation
- Resumen Inmunologia AcoDocument135 pagesResumen Inmunologia AcoManuel González AlayesPas encore d'évaluation
- Estructura y función del TCR y BCR en linfocitos T y BDocument6 pagesEstructura y función del TCR y BCR en linfocitos T y BEsther MoralesPas encore d'évaluation
- Anatomia y Elementos CelularesDocument7 pagesAnatomia y Elementos CelularesPilar del Valle Pico GutierrezPas encore d'évaluation
- Clase 10. ONTOGENIA DE LAS CÉLULAS LINFOIDESDocument6 pagesClase 10. ONTOGENIA DE LAS CÉLULAS LINFOIDESKevin VelaPas encore d'évaluation
- Expoooo Receptores para Antígeno e InmunoglobulinaDocument25 pagesExpoooo Receptores para Antígeno e InmunoglobulinaCdcgs PWinPas encore d'évaluation
- Cap 10Document7 pagesCap 10Natalia Espíndola BalderasPas encore d'évaluation
- Cuarto y Último SeminarioDocument5 pagesCuarto y Último SeminarioAle CarmonaPas encore d'évaluation
- Parcial 3 InmunoDocument15 pagesParcial 3 InmunoROGER DIAZPas encore d'évaluation
- Autoevaluaciones Examen Final InmunoDocument31 pagesAutoevaluaciones Examen Final InmunoRovian RosaliaPas encore d'évaluation
- Cuestionario SEMANA 3Document11 pagesCuestionario SEMANA 3Ericksen SalazarPas encore d'évaluation
- Linf T 1Document69 pagesLinf T 1Alex David Sigueñas NoriegaPas encore d'évaluation
- Rpta Inmune Humoral. Noviembre 2013Document51 pagesRpta Inmune Humoral. Noviembre 2013Jose Marco Muñoz BocanegraPas encore d'évaluation
- Mecanismo de Rechazo CelularDocument6 pagesMecanismo de Rechazo Celular11-MH-HU-AMELIA DANITZA CACERES AMAROPas encore d'évaluation
- El sistema inmunitario: células y componentes claveDocument13 pagesEl sistema inmunitario: células y componentes claveGabriel Hernandez MartinPas encore d'évaluation
- Nucleo 2 Cooperacion TBDocument57 pagesNucleo 2 Cooperacion TBLUCIA AGUILARPas encore d'évaluation
- Resumen Segundo Parcial (Marilin)Document21 pagesResumen Segundo Parcial (Marilin)Nico Alister CortésPas encore d'évaluation
- Conociendo Mi Grupo Sanguíneo.Document9 pagesConociendo Mi Grupo Sanguíneo.Deyvis Andres Nieves FloresPas encore d'évaluation
- Preguntas de Inmunología 1,2,3,4Document38 pagesPreguntas de Inmunología 1,2,3,4Mishelle CastroPas encore d'évaluation
- Maduración de Linfocitos TDocument41 pagesMaduración de Linfocitos TVicToRia50% (2)
- Rol MHC presentación antigénicaDocument3 pagesRol MHC presentación antigénicaLeandroCayaoPas encore d'évaluation
- RESPUESTA INMUNE CELULARDocument28 pagesRESPUESTA INMUNE CELULARraziel boxPas encore d'évaluation
- Activacion de Los Linfoncitos T.Document6 pagesActivacion de Los Linfoncitos T.Lis Juli MenRePas encore d'évaluation
- Activacion Del Linfocito TDocument8 pagesActivacion Del Linfocito TMANUEL ALBERTO VERA PIMINCHUMOPas encore d'évaluation
- Republica Bolivariana de Venezuela Medico QuirurgicoDocument26 pagesRepublica Bolivariana de Venezuela Medico QuirurgicoBarbara DiazPas encore d'évaluation
- Notas Rápidas PDFDocument2 pagesNotas Rápidas PDFMónica Andrea Quispe RojasPas encore d'évaluation
- 1) ¿Cuántas Formas de Denominar Se Conoce? 2) ¿Cómo Está Constituido El LCR?Document2 pages1) ¿Cuántas Formas de Denominar Se Conoce? 2) ¿Cómo Está Constituido El LCR?AimméMirandaMoralesPas encore d'évaluation
- Evaluación - TejidosDocument3 pagesEvaluación - TejidosCristina Moreno LeuroPas encore d'évaluation
- Guía 1 Biología 6° Generalidades de La Célula y Teoría Celular.Document10 pagesGuía 1 Biología 6° Generalidades de La Célula y Teoría Celular.wilson Andres Cardenas MejiaPas encore d'évaluation
- Triptico Del CorazonDocument2 pagesTriptico Del CorazonCarlitos Valentin Laureano100% (3)
- Ahora Dialoga Con Tus Compañeros de Grupo y Respondan Las Siguientes PreguntasDocument3 pagesAhora Dialoga Con Tus Compañeros de Grupo y Respondan Las Siguientes PreguntasAlexander Gino Alva LeónPas encore d'évaluation
- Xilema y FloemaDocument10 pagesXilema y FloemaJherson Guillermo Bonifacio EspinozaPas encore d'évaluation
- Rsquema de PleuraDocument1 pageRsquema de PleuraJuan R. AntunezPas encore d'évaluation
- Piel anatomía histología fisiología preguntas bancoDocument103 pagesPiel anatomía histología fisiología preguntas bancoJhonny HuancaPas encore d'évaluation
- Construye Un Mapa Conceptual de Los Tipos de Neuronas Explicando Sus Funciones.Document1 pageConstruye Un Mapa Conceptual de Los Tipos de Neuronas Explicando Sus Funciones.GuadalupePas encore d'évaluation
- Masoterapia terapéuticaDocument10 pagesMasoterapia terapéuticaCesar Alejandro HerreraPas encore d'évaluation
- Resumen Vision, Audicion EtcDocument11 pagesResumen Vision, Audicion EtcRuth Esther Nuñez MaldonadoPas encore d'évaluation
- Anatomía de OídoDocument5 pagesAnatomía de OídoMONTSERRATH RAMIREZ MORALESPas encore d'évaluation
- Manual Neurología y Neurocirugía 6a ediciónDocument104 pagesManual Neurología y Neurocirugía 6a ediciónPavel Rojas OlivaresPas encore d'évaluation
- Monografias Traumatologia N2 Sergio CruzDocument109 pagesMonografias Traumatologia N2 Sergio Cruzsergio cruzPas encore d'évaluation
- TESIS CINTHYA VICTORIA CANTOR CUEVAS Estrias 4Document76 pagesTESIS CINTHYA VICTORIA CANTOR CUEVAS Estrias 4Alejandro Lenis CalzadaPas encore d'évaluation
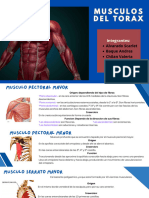
- Musculos Del ToraxDocument7 pagesMusculos Del ToraxValeria kPas encore d'évaluation
- Musculos InfrahioideosDocument9 pagesMusculos InfrahioideosElii DíazPas encore d'évaluation
- Caso clínico de artritis reactiva en adolescenteDocument11 pagesCaso clínico de artritis reactiva en adolescenteHector Crhistian Vilca HuisaPas encore d'évaluation
- Texto FgekDocument39 pagesTexto FgekAngela PacoPas encore d'évaluation
- Semiologia de Las Glandulas Salivales y Principales Alteraciones PDFDocument11 pagesSemiologia de Las Glandulas Salivales y Principales Alteraciones PDFRuddy Frank67% (3)
- Explicación Dinámica de La Contracción MuscularDocument18 pagesExplicación Dinámica de La Contracción MuscularAlicia Lopez OrtizPas encore d'évaluation
- EsplenectomíaDocument2 pagesEsplenectomíaElizabethPas encore d'évaluation
- 01 m2t1 Experto en CitologiaDocument20 pages01 m2t1 Experto en CitologiaFelvir RivasPas encore d'évaluation