Académique Documents
Professionnel Documents
Culture Documents
Biología Celular
Transféré par
David AldamaCopyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Biología Celular
Transféré par
David AldamaDroits d'auteur :
Formats disponibles
BIOLOGA CELULAR
Csar Eduardo Montalvo Arenas
11/09/03
LA CLULA.
La clula es la unidad morfolgica y funcional fundamental
de todo ser vivo. La Teora celular considera que: todas las
formas de vida existentes en nuestro planeta (seres
unicelulares, animales y vegetales) estn constituidas por
clulas y sus productos de secrecin; que cualquier clula se
origina de otra clula preexistente y que cada clula posee
vida propia, independiente de su integracin a un organismo
pluricelular.
Para llegar a este enunciado, debieron transcurrir varios
cientos de aos para que se le descubriera y fuera aceptado
por la comunidad cientfica del mundo. El conocimiento de
la existencia de las clulas slo fue posible que se llevara a
cabo, cuando se desarrollaron los primeros instrumentos
pticos que permitieron ampliar la imagen de la casi
totalidad de ellas, porque al poseer dimensiones tan
pequeas era imposible realizar la observacin de las clulas
a simple vista.
Los primeros microscopios simples y compuestos que se
empezaron a fabricar a partir del siglo XVI facilitaron la
tarea de los investigadores cientficos de examinar y
observar seres vivos pequeos, tejidos animales y vegetales,
ciertos fluidos animales como la sangre, el semen y
animculos de aguas estancadas.
HALLAZGOS QUE PERMITIERON ENUNCIAR LA
TEORA CELULAR.
Los primeros microscopios compuestos fueron construidos
por Francis y Zacharias Janssens, en 1590. Permitan la
amplificacin de las imgenes en un rango de 10 a 30
dimetros. A travs de ellos se podan observar insectos
pequeos.
observaciones presentados en la Real Sociedad de Londres
abarcaban desde bacterias (cocos, bacilos y espirilos) hasta
espermatozoides de varias especies animales; el movimiento
de las clulas de la sangre en capilares de tejidos
translcidos de ranas y conejos. Pudo describir la diferencia
entre los eritrocitos de peces y anfibios, ovalados y con un
corpsculo central (ncleo) y los de los mamferos redondos
y sin el corpsculo central (anucleados).
Figura Biol. Cel.1. Imgenes dibujadas por Leeuwenhoek de las
estructuras observadas mediante su microscopio. (pag. 5 Mol. Cell. Biol.
Darnell, 3 ed. 1995).
Robert Hooke (1635 - 1703), arquitecto y cientfico ingls,
fue el impulsor del uso de los microscopios para los
bilogos ingleses. El mismo fue un excelente constructor de
microscopios. Una de sus observaciones ms trascendentes
fue el que realiz en secciones delgadas de la corteza del
rbol del corcho. Describi que aquellas estaban perforadas
y porosas muy semejantes a un panal. A este conjunto de
estructuras formadas por paredes y espacios les denomin
clulas o celdillas. Se considera a Robert Hooke como el
investigador que utiliz por primera vez la palabra clula.
Marcelo Malpighi, cientfico italiano que vivi entre 1628 y
1694,
realiz las primeras descripciones de la
microanatoma animal y vegetal. Fue el primero en describir
los capilares sanguneos; as ayud a corroborar los estudios
que sobre la circulacin sangunea haba expuesto aos atrs
el fisilogo ingls William Harvey.
Malpighi fue el primer investigador que observ secciones
delgadas de tejidos animales y vegetales y tambin realiz
estudios sobre el desarrollo embriolgico del pollo. En sus
ltimos aos de vida Malpighi se dedico especialmente al
estudio microscpico de las plantas.
Otro microscopista, pionero en este campo, fue el holands
Antonie van Leeuwenhoek (1632-1723), experto tallador de
lentes; lleg a construir microscopios con aumentos que, se
afirma, alcanzaban hasta 300 dimetros. Sus hallazgos y
Figura Biol.. Cel. 2. Imagen de las paredes celulares que Hooke observ
en la corteza del corcho (alcornoque) a las cuales denomin celdillas.
(pag. 6. Mol. Cell Biol. Darnell, 3 ed. 1995).
Otro investigador ingls, Nehemiah Grew (1641 - 1712),
junto con Malpighi, publicaron una serie de informes donde
describieron imgenes microscpicas de secciones de flores,
races y tallos los cuales estaban constituidos por clulas.
En 1824, Ren Dutrochet (1776-1847) lleg a la conclusin
que todos los tejidos animales y vegetales estn integrados
por agregados de clulas globulares.
En 1831, el ingls Robert Brown ( 1773-1858) observ que
las clulas vegetales de la epidermis de las hojas, los granos
de polen y clulas del estigma contenan unos corpsculos
constantes a los que denomin ncleos, de esta manera
este trmino se incorpor al lxico de la biologa.
Un cientfico polaco, Johannes Purkinje (1787- 1869) le
puso el nombre de protoplasma al contenido de las
clulas.
Un alemn, Mathias J. Schleiden (1804-1881), contino los
estudios sobre el ncleo en vegetales, hechos por Robert
Brown, y el mismo descubri el nucleolo. Se atribuye a
Schleiden el reconocimiento de la individualidad
morfolgica y funcional de las clulas vegetales y su
comportamiento integral en la planta.
Otro alemn, Theodor Schwann (1810-1882), realiz
estudios en tejidos conjuntivos (cartlago y hueso).
Consider que los animales estn constituidos por clulas y
por el producto que ellas sintetizan y secretan.
A Schleiden y a Schwann se les consideran los cientficos
que plantearon uno de los principios de la teora celular al
afirmar que, todos los seres vivos vegetales y animales
pluricelulares estn integrados por clulas y sus productos
de secrecin. Schwann fue el que utiliz el trmino
metabolismo para explicar las actividades de la clula.
componentes citoplasmticos como los organelos
membranosos y no membranosos, el ncleo, el
citoesqueleto, productos de sntesis y secrecin celular, etc.
Por lo tanto podemos definir a la clula como:
La unidad morfolgica, funcional y gentica de todo ser
vivo (perteneciente a los reinos monera, protista, fungi,
plantae y animalia) y sus productos que sintetizan y
secretan.
CONSTITUCIN CELULAR DE LOS SERES VIVOS.
Los seres vivos pueden estar formados por una sola clula;
denominados unicelulares o constituidos por ms de una
clula; en este caso se le conoce como multicelulares o
pluricelulares.
Los seres unicelulares son bioqumica y funcionalmente
complejos pues tienen la capacidad de realizar todas las
funciones biolgicas relacionadas con su metabolismo en
general, y de generar otras clulas (hijas) con las mismas
caractersticas morfolgicas y funcionales.
Los seres pluricelulares, en cambio, al estar integrados por
varias o muchas clulas de morfologa y funciones
especficas o restringidas, requieren de la cooperacin de
otras clulas que forman parte del organismo pluricelular
para conferirle a ste, la capacidad de cumplir a cabalidad su
metabolismo y su funcin de reproduccin.
Un patlogo alemn, Rudolf Virchow (1821-1902) reconoci
que las enfermedades tienen un sustento celular. Sus
estudios le permitieron enunciar la base celular de la
continuidad de la vida al afirmar que omnis cellula e
cellula(todas las clulas se originan de otras clulas).
A fines del siglo XIX y durante todo el siglo XX el
microscopio fotnico alcanz su mximo desarrollo y
perfeccionamiento (microscopio de contraste de fases, de
campo oscuro, de contraste interferencial diferencial, el de
polarizacin y el de fluorescencia que utiliza radiacin
ultravioleta) aunado al empleo de una serie de tcnicas
histolgicas, histoqumicas, de inmunohistoqumicas y de
inmunofluorescencia facilit el estudio de los componentes
celulares y varias de sus funciones. A partir de la dcada de
los aos 40, el invento del microscopio electrnico, su
desarrollo y perfeccionamiento ampli de manera inusitada
el panorama morfolgico de las clulas.
Las imgenes aumentadas varios decenas de veces de las
imgenes que ofrece un microscopio fotnico, redundaron
en el conocimiento de estructuras ultramicroscpicas de
todos los componentes celulares: membrana celular,
a)
b)
Figura Biol. Cel. 3. a) seres unicelulares, b) un ser pluricelular.
CLASIFICACIN DE LOS SERES VIVOS POR SU
ESTRUCTURA CELULAR.
El desarrollo del microscopio electrnico permiti a la
comunidad cientfica, a partir de 1950 aproximadamente,
observar imgenes de las clulas con varios miles de
dimetros y lograr describir una serie de detalles de los
componentes de las clulas.
Estas observaciones corroboraron la clasificacin que en
1937, Chatton, un cientfico francs hizo para diferenciar a
las clulas en procariotas (clulas sin ncleo) y eucariotas
(clulas con ncleos) y demostrar que adems de la carencia
o presencia del ncleo las clulas de los seres vivos estn
constituidas por una serie de estructuras que guardan
diferencias y semejanzas en sus estructuras morfolgicas y
caractersticas funcionales. Las evidencias cientficas
recogidas hasta el momento indican que las clulas
procariotas existieron mucho tiempo antes que las
eucariotas. stas se formaron a partir de clulas procariotas
a travs de una serie de procesos evolutivos que duraron
millones de aos.
CARACTERSTICAS
QUE
IDENTIFICAN
Y
DIFERENCIAN A LAS CLULAS PROCARIOTAS DE
LAS EUCARIOTAS.
Rasgos estructurales: Ambos linajes celulares poseen una
membrana plasmtica o plasmalema de estructura qumica
similar en sus principales componentes moleculares, que
acta como una barrera permeable selectiva entre el interior
de la clula y el microambiente exterior. Las clulas
procariotas y eucariotas (de hongos y plantas) poseen
adems una pared celular rgida o semirgida, que protege
al plasmalema y al citoplasma celular de agresiones del
exterior, aunque sus componentes qumicos suelen ser
diferentes.
Identidad gentica: Ambas clulas poseen una regin donde
se acumula el material gentico, casi siempre rodeada de
citoplasma. En la clula procariota el material gentico,
denominado nucleoide, se encuentra libre en el citoplasma,
no est rodeado por una membrana, en cambio en la clula
eucariota este material, denominado ncleo esta rodeado
por una membrana o cubierta nuclear.
Esta diferencia en el contenido gentico permite la
utilizacin de los trminos procariota (Pro = antes; carion =
nuez, ncleo) y eucariota (Eu = verdadero; carion = nuez,
ncleo).
La cantidad y disposicin del material gentico o cido
desoxirribonucleico (DNA) es diferenciable en las clulas
procariotas y eucariotas. En las procariotas el DNA alcanza
una longitud entre 0.25 mm y 3 mm, en las clulas
eucariotas (con excepcin de las levaduras cuyo DNA mide
aproximadamente 4.6 mm) la cadena de DNA alcanza
longitudes del orden de varios centmetros, inclusive los
seres unicelulares.
Figura Biol. Cel. 4. a) clula eucariota, b) clula procariota.
Ambos tipos de clulas organizan el DNA en forma de
cromosomas; las procariotas poseen un cromosoma
circular carente de protenas organizadoras, mientras que
las eucariotas ordenan su material gentico en cadenas
cromosmicas (cromosomas) que contienen DNA y
protenas.
Divisin celular: Las clulas eucariotas se reproducen
mediante dos mecanismos complejos, las clulas somticas
mediante mitosis y las clulas germinales o sexuales a
travs de la meiosis. En ambos procesos los cromosomas
duplican su material de DNA, se condensan en cromosomas
compactos y visibles para que, a continuacin, stos sean
separados por un complicado sistema de microtbulos (el
huso acromtico). En las procariotas, el cromosoma no se
condensa y tampoco se forma el huso, el DNA se duplica y
luego las dos copias se separan por fisin debido al
crecimiento de una membrana celular que se interpone entre
las dos cadenas de DNA.
Organelos citoplasmticos: El contenido del citoplasma de
ambas clulas tambin es diferente. En las clulas eucariotas
existe una gran diversidad de organelos, varios constituidos
por membranas (mitocondrias, retculo endoplsmico liso y
rugoso, complejo o aparato de Golgi, lisosomas,
peroxisomas, grnulos de secrecin; cloroplastos y vacuolas
en clulas vegetales) y otros carentes de ellas (centrolos,
microtbulos, filamentos del citoesqueleto, proteasomas,
ribosomas). En las clulas procariotas existen escasos
organelos; de los membranosos se puede mencionar a los
mesosomas, simples invaginaciones de la membrana celular
y las membranas fotosintetizadoras de las algas verdiazules
o cianobacterias. Tambin contienen ribosomas; stos son
de menor tamao que aquellos de las eucariotas. En ambos
casos desarrollan la funcin de ensamblar aminocidos para
constituir pptidos y protenas.
Cilios y flagelos: Las clulas procariotas poseen un
mecanismo bastante simple de locomocin. El flagelo,
delgado filamento protenico de flagelina, genera
movimientos de rotacin que, en un medio lquido, impulsa
la bacteria hacia adelante. En las eucariotas por ejemplo, las
que pertenecen al reino protistas, poseen flagelos y cilios, de
estructura protenica (tubulina) ms compleja. Los
espermatozoides de los seres pluricelulares, tambin son
clulas flageladas.
Tamao. Las clulas procariotas miden entre 1 a 10
micrmetros, aproximadamente, de dimetro o longitud. Las
clulas eucariotas, generalmente miden de 10 a 100
micrmetros de dimetro o longitud. Existen excepciones,
por ejemplo las neuronas de la capa de los granos del
cerebelo, cuyo cuerpo celular mide entre 4 a 5 micrmetros
de dimetro o los eritrocitos de mamferos, cuyos tamaos
pueden ser de 5 a 8 micrmetros o las fibras musculares
estriadas esquelticas que suelen medir varios centmetros
de longitud.
CARACTERSTICAS ESTRUCTURALES DE UNA
CLULA EUCARIOTA ANIMAL.
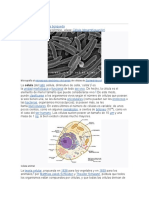
La clula eucariota animal esta constituida por:
Membrana celular o plasmalema. Es la cubierta externa de
la clula, est constituida por lpidos, protenas y
carbohidratos. Forma una barrera permeable selectiva entre
el denominado protoplasma (citoplasma + ncleo) y el
microambiente externo.
Citoplasma. Es la porcin de la clula comprendida entre la
membrana celular y el ncleo. Es una sustancia lquida
constituida principalmente por agua (citosol), en ella se
suspenden o disuelven sustancias orgnicas e inorgnicas.
En el citosol se encuentran una serie de estructuras
microscpicas denominados organelos que desarrollan,
cada uno de ellos, funciones especficas. Los organelos
estn formados por membranas celulares o carecen de ellas.
La forma y el tamao de las clulas estn sustentados
principalmente por una serie de organelos no membranosos,
generalmente de aspecto fibrilar, que integran el
denominado citoesqueleto.
Ncleo. Es una estructura esfrica, ovalada, alargada o
lobulada, rodeada por una doble membrana celular
(cubierta nuclear) que alberga, en su interior, el material
gentico de las clulas (DNA y protenas), organizado en
finos filamentos denominados cromosomas y el cido
ribonucleico (RNA) constituyente orgnico, directamente
responsable de la sntesis de protenas.
Figura Biol.Cel. 5. Componentes de una clula eucariota.
TAMAO DE LAS CLULAS EUCARIOTAS.
Las clulas poseen diversos tamaos; generalmente
microscpicos. En la gran mayora de los casos nicamente
pueden observarse con la ayuda de los microscopios
(fotnicos y electrnicos), salvo ciertas excepciones como
los ovocitos de ciertos vertebrados (peces, anfibios, reptiles
y aves).
Dependiendo del reino al que pertenezcan pueden medir
desde 1.0 a 5.0 m (levaduras) hasta 120 m (ovocitos de
mamferos o cuerpos celulares de neuronas). Las clulas
eucariotas suelen medir, en promedio, entre 10 a 30 m.
Existen causas y caractersticas celulares para que las
clulas posean tamaos microscpicos:
El ncleo de la clula contiene la informacin gentica
necesaria para transcribir rdenes, a travs del RNA
mensajero, con la finalidad de sintetizar ciertas
cantidades de determinadas protenas. Cuanto mayor
sea la cantidad de citoplasma existente en una clula,
ser insuficiente la sntesis de protenas para el
adecuado funcionamiento de la misma. La existencia de
clulas binucleadas (clulas del hgado) o
multinucleadas
(como las fibras
musculares
esquelticas o los osteoclastos), todas ellas con
abundante cantidad de citoplasma son la prueba ms
fehaciente de lo afirmado.
Conforme el tamao de la clula aumenta, la
proporcin entre la superficie y el volumen se
desequilibra (disminuye). Esto afecta la capacidad de la
clula de intercambiar sustancias con el microambiente
que la rodea y, la superficie no sera la adecuada para
un suficiente intercambio de gases y nutrimentos que
requiere el realizar un metabolismo eficiente.
Las unidades de medidas empleadas para conocer el tamao
de las clulas y de sus componentes microscpicos se hacen
en dimensiones lineales que corresponden a fracciones de la
milsima parte de un metro, el milmetro. Las unidades
lineales ms utilizadas son la micra o micrmetro ( m); el
nanmetro (nm) y el Angstrom ().
La micra o micrmetro equivale a la milsima parte de
un milmetro 10-6 de metro. Un nanmetro equivale a
la milsima parte de una micra o micrmetro o 10-9
de metro y el Angstrom equivale a la dcima parte de
un nanmetro o 10-10 de metro.
Mediante el microscopio fotnico nicamente se pueden
medir clulas y componentes de ella, empleando la micra o
el micrmetro, en cambio, con el microscopio electrnico es
posible utilizar las tres unidades antes mencionadas.
Formas celulares. Las clulas de los organismos
unicelulares o pluricelulares poseen una gran diversidad de
formas. En el caso de los seres animales pluricelulares y de
manera especfica la de los mamferos (incluida la especie
humana) suelen ser:
Redondas o esfricas. Por ejemplo: ovocitos, leucocitos en
el plasma sanguneo.
Aplanadas o escamosas: Clulas en las que el largo y el
ancho predominan sobre el espesor o grosor. Ejemplos: las
de los alvolos pulmonares, de la hoja parietal de la cpsula
de Bowman, las clulas endoteliales y mesoteliales, las
clulas del estrato crneo o queratinizado de la epidermis
(piel) de la vagina, cavidad bucal, etc.
Clulas cbicas. En ellas las dimensiones de largo, ancho y
grosor, son sensiblemente similares; por ejemplo: las clulas
que forman los tbulos renales, las vesculas o folculos
tiroideos, los plexos coroideos, las glndulas sudorferas.
Clulas cilndricas o columnares. Son clulas en las que el
largo o altura tiene la dimensin mayor, en cambio el ancho
y el grosor son semejantes. Se les observa en el epitelio
secretor del estmago y en el de absorcin intestinal, en el
tero, en los conductos menores de las glndulas, en el
epitelio de la vescula biliar.
Clulas polidricas. Poseen varias superficies de
dimensiones ms o menos similares, por ejemplo los
hepatocitos (clulas del hgado), las clulas de la corteza
suprarrenal, el estrato espinoso de la epidermis, las clulas
de Leydig del parnquima testicular endocrino.
Clulas discoidales bicncavas: Muestran un aspecto de
disco con sus dos superficies cncavas; es la forma que
poseen los eritrocitos de la gran mayora de los mamferos.
Clulas ovaladas. El ejemplo ms caracterstico son los
eritrocitos (nucleados) de varios vertebrados: Anfibios,
reptiles y aves. En mamferos, las especies de camlidos: los
camellos, dromedarios, llamas, alpacas etc., poseen
eritrocitos anucleados ovalados. Otros ejemplos de clulas
que exhiben un contorno ovalado son los mastocitos o
clulas cebadas y las clulas plasmticas.
Figura Biol. Cel. 6. Diferentes formas y tamaos de clulas eucariotas.
Clulas cilndricas alargadas. Se le conocen tambin como
fibras. Por ejemplo las clulas o fibras musculares
esquelticas. Suelen medir, en ciertos casos, hasta varios
centmetros de longitud.
Clulas fusiformes. Tienen forma de huso, son
ligeramente alargadas. Sus extremos terminan en punta,
mientras que el centro de la clula es ms engrosado.
Muestran esta forma las clulas fibras musculares lisas y
los fibroblastos.
Clulas estrelladas. Son clulas que poseen un cuerpo
polidrico del cual emergen una serie de prolongaciones
citoplasmticas que, en conjunto, le confieren una forma
estrellada a la clula. As se observan la mayora de las
neuronas, las clulas gliales y las mesenquimatosas o
embrionarias.
PROPIEDADES
CLULAS
FUNDAMENTALES
DE
LAS
A) Organizacin y complejidad de las clulas: Las clulas
se organizan bioqumicamente de tal manera que, en ellas,
se expresa la vida. Los niveles de organizacin abarcan
desde la unin de tomos, ensamble de molculas simples o
compuestas las cuales constituyen complejos polimricos,
que siguiendo un patrn concordante y predecible se
organizan en organelos y finalmente en una clula.
B) Existencia de un cdigo gentico y capacidad de
utilizarlo: Las clulas se originan de otras clulas a partir de
la informacin contenida en un cdigo gentico. Esta
informacin, dispuesta en forma de genes, se encuentra
empaquetada en los cromosomas dentro del ncleo. Los
genes son unidades moleculares de DNA que contienen la
informacin necesaria para: generar estructuras celulares,
poner en marcha y mantener las funciones de la clula y
lograr reproducirse as mismas.
C) Capacidad de reproducirse a s mismas. Una clula al
dividirse origina dos clulas hijas (nuevas), esto significa
que el contenido de DNA de la clula madre se hereda y
distribuye a las dos clulas hijas. Antes de la divisin el
DNA se duplica fielmente y cada clula hija recibe una
dotacin completa e idntica de la informacin gentica.
Casi en la totalidad de los casos, las clulas hijas tambin
reciben cantidades similares de contenido citoplasmtico,
excepto los ovocitos de mamferos, uno de los cuales
durante la meiosis, retiene casi todo el citoplasma aunque
solo herede la mitad del material gentico.
D) Las clulas captan y utilizan energa. Las diversas
funciones que realiza una clula requieren necesariamente
del consumo de energa. En las clulas animales la energa
se obtiene a partir de incorporar carbohidratos a la economa
celular, la glucosa es el azcar que se emplea para tal fin. La
descomposicin de la glucosa (energa qumica) dentro de la
clula resulta en transformacin en un compuesto altamente
energtico, la adenosinatrifosfato (ATP) que almacena la
energa y la libera rpidamente para producir trabajo celular.
Este proceso de generar y almacenar energa se produce en
las mitocondrias, organelos membranosos existentes en
todas las clulas eucariotas.
E) Realizacin de mltiples reacciones qumicas. Dentro
de la clula se producen miles de reacciones qumicas con
la finalidad que ella cumpla las diversas funciones. La
mayora de ellas se producen a travs de enzimas que
aceleran los procesos qumicos. A este conjunto de procesos
qumicos se les conoce como metabolismo. El metabolismo
celular consta de dos procesos, el anabolismo cuya
finalidad es sintetizar sustancias estructurales, funcionales
y de exportacin de la clula y el catabolismo, proceso
encargado de descomponer sustancias y cuyos productos no
tiles sean eliminados de las clulas.
F) Produccin de actividad motriz. Las clulas requieren
de transportar sustancias dentro de ellas para su actividad
metablica o para que ella misma pueda desplazarse en su
microambiente o generar movimientos como la de cilios y
flagelos. Esta capacidad de movimiento se produce por la
interaccin de molculas protenicas que intervienen como
pequeos motores para la actividad mecnica de las
clulas. Las clulas que muestran un alto grado de esta
funcin, la contractibilidad son las fibras musculares de
nuestro organismo.
G) Capacidad para captar estmulos y generar una
respuesta. Una serie de clulas responden a diversidad de
estmulos. Los organismos unicelulares como los ciliados,
flagelados o aquellos que emiten seudpodos (paramecios,
euglenas y amibas, respectivamente) ante determinados
estmulos se desplazan para evitar un obstculo puesto en su
recorrido o se acercan a un lugar donde existen alimentos o
se alejan cuando sienten que una sustancia puede
ocasionarles dao.
En los seres pluricelulares, existen clulas que han
desarrollado una gran capacidad para captar estmulos
(irritabilidad) y generar una respuesta efectora que suele
ser motriz o secretora. En la membrana celular existen una
serie de receptores que interactan con el microambiente
que las rodea. Los estmulos son hormonales, por
electrolitos, neurotransmisores, factores de crecimiento o
estmulos fsicos y qumicos que provienen del medio
externo (tacto, luz, sonido, sabor, olor). En ciertos casos las
clulas responden a los estmulos alterando su metabolismo,
iniciando la divisin celular, sintetizando protenas
especficas para diferenciarse o a veces para dar trmino a
su capacidad vital. Un ejemplo de clulas del organismo
animal que han desarrollado en grado extremo, esta
capacidad de captar estmulos y elaborar una respuesta son
las neuronas.
H) Capacidad de autorregulacin. El complejo sistema
estructural y funcional que posee una clula le permite, a su
vez, regular su capacidad de sntesis o destruccin de
sustancias, de reproducirse o de generar una respuesta
motora o secretora. La actividad y la informacin contenida
en los cidos nucleicos (DNA y RNA) permiten que en cada
clula existan momentos precisos y acciones espontneas
que facilitan la secuencia de pasos para alcanzar un
propsito funcional determinado.
MEMBRANAS CELULARES.
Las clulas de todo organismo vivo (procariotas y
eucariotas) estn limitadas por una cubierta denominada
membrana celular, membrana plasmtica o plasmalema.
Forma una barrera permeable, de manera selectiva, entre el
citoplasma, el ncleo y el medio externo. La existencia de
la membrana celular es la caracterstica ms importante en
la organizacin celular. Una serie de organelos ( retculo
endoplsmico, el complejo de Golgi, mitocondrias,
lisosomas, etc.) y la cubierta del ncleo tambin estn
constituidos por membranas que guardan semejanza
estructural y funcional con el plasmalema celular.
Las membranas celulares cumplen varias funciones:
a) Conservan la integridad de la clula y de los
componentes citoplasmticos y nucleares.
Las
membranas celulares son lminas que rodean a los
compartimentos celulares. El plasmalema rodea a todo el
contenido protoplasmtico. La cubierta nuclear y las
membranas de organelos citoplasmticos circundan e
individualizan una serie de espacios internos en los cuales se
generan mltiples procesos y actividades especializadas, sin
que existan interferencias externas. Estos compartimentos
membranosos albergan una serie de sustancias lquidas o en
suspensin que no deben mezclarse.
b) Constituyen barreras permeables selectivas. Las
membranas son medios que facilitan la comunicacin entre
un espacio y otro de manera selectiva pero, a su vez impiden
el libre intercambio de sustancias entre el medio externo y
entre los microambientes internos. El plasmalema garantiza
que las sustancias necesarias penetren hacia el citoplasma y,
las del producto del anabolismo y del catabolismo salgan de
la clula. Todo ello permite que el citoplasma se conserve en
condiciones de homeostasis, y se produzcan las reacciones
bioqumicas necesarias para mantener la vida.
Homeostasis. Estado de equilibrio de una clula o de un
organismo pluricelular con relacin a sus componentes
morfolgicos y funcionales y la regulacin estable entre el
anabolismo y el catabolismo.
c) Regular las interacciones entre las clulas. La
membrana celular interviene como mediadora entre las
clulas de un ser vivo pluricelular. El plasmalema permite a
las clulas vecinas adherirse entre ellas, reconocerse
mutuamente e intercambiar sustancias.
d) Transporte de solutos. La membrana celular est
constituida para transportar sustancias del medio externo al
citoplasmtico y tambin en sentido contrario. Desde una
regin donde la concentracin de solutos es menor a otra
regin donde los solutos estn en mayor concentracin y
viceversa. Esta actividad se produce, en ciertos casos,
mediante un transporte pasivo a travs del transporte
activo es decir por cambios estructurales o bioqumicos de
la membrana (endocitosis y exocitosis, utilizacin de
molculas acarreadoras que se traducen en la necesidad de
gastar grandes cantidades de energa metablica).
El transporte a travs del plasmalema le permite a la clula
acumular azucares, lpidos y aminocidos como fuentes de
reserva energtica y la sntesis de macromolculas. De la
misma manera suele separar iones con cargas elctricas
opuestas e integrar gradientes de iones en el citoplasma tal
como sucede en las clulas nerviosas (neuronas) y en las
fibras musculares.
e) Respuesta a seales externas o transduccin de
seales. La membrana plasmtica acta como una barrera
sensible a estmulos provenientes de su microambiente
externo. Las membranas celulares poseen una serie de
molculas integradas a ellas conocidos como receptores que
se combinan con ligandos, molculas especficas
extracelulares de estructura qumica complementarias.
Los diferentes tipos de clulas poseen distintos tipos de
ligandos en sus membranas, por lo tanto pueden reconocer y
responder a diversos estmulos o seales. Los ligandos de
actividad ms frecuentes son los neurotransmisores, las
hormonas y los factores de crecimiento, stos se unen a los
receptores pero no atraviesan la membrana.
La unin del ligando con el receptor externo provoca que la
clula genere una seal que estimula o inhibe alguna
funcin especfica de la clula, generalmente relacionada
con una respuesta de secrecin o motora. Por ejemplo que la
clula libere sustancias inflamatorias o secrete enzimas para
degradar tejido seo; se prepare para iniciar un proceso
mittico (divisin celular) o para almacenar glucosa en
forma de glucgeno, que inicie procesos de contraccin
molecular y pueda desplazarse a otros lugares o generar
movimientos de otras clulas o tejidos, como los realizados
por las fibras musculares o recibir la seal que debe dejar de
funcionar y morirse (apoptosis).
Los receptores pueden reconocer ligandos de clulas
extraas al organismo o de clulas propias pero enfermas o
anormales.
f) Poseen lugares para realizar actividades bioqumicas.
Las membranas celulares intervienen como un medio para
coordinar y organizar actividades celulares. Las membranas
celulares le proporcionan a la clula una extensa superficie
de soporte para la secuencia adecuada de procesos
enzimticos y la obtencin de una reaccin metablica. La
mayor cantidad y calidad de las reacciones bioqumicas
celulares tienen su apoyo en la presencia de organelos
celulares membranosos.
g) Transformacin de la energa. Las membranas
celulares intervienen en los procesos de transformacin de
una energa en otra (transduccin de energa). Un ejemplo
de esta accin es la que se produce en las plantas a travs
del proceso de la fotosntesis en la cual los pigmentos de
clorofila asociados a membranas de los cloroplastos,
absorben energa luminosa del sol, la transforman en energa
qumica y la almacenan en forma de carbohidratos:
azucares, almidn, pectina etc. Las membranas de las
mitocondrias intervienen en la transformacin de la energa
qumica de azucares y lpidos a adenosinatrifosfato (ATP).
ESTRUCTURA DE LAS MEMBRANAS CELULARES.
Estructura morfolgica. Las membranas celulares en
general y el plasmalema, de manera particular, no se pueden
observar con el microscopio fotnico. Las imgenes
obtenidas mediante el microscopio electrnico, las muestran
como hojas trilaminares muy finas, constituidas por tres
zonas, dos electrondensas, una externa y otra interna,
separadas por una zona electronlcida. En promedio las
membranas miden 7.5 nanmetros de grosor, cada una de
las tres zonas miden 2.5 nm aproximadamente.
Biol. Cel. 7. Imagen de la membrana celular observada a travs del
microscopio electrnico.
Estructura bioqumica. En 1899, E. Overton reconoci que
los lmites existentes en las clulas vegetales y animales
estaban impregnados de lpidos. Su teora se basaba en
que despus de poner en contacto el contorno celular de las
clulas vegetales con mas de 500 sustancias de diverso tipo,
observ que los compuestos solubles en solventes orgnicos
se introducan a las clulas con mayor rapidez que los
compuestos solubles en agua; dedujo que las sustancias
liposolubles se disolvan en los componentes lipdicos de la
membrana y de esa manera tenan mayor y ms rpida
accesibilidad al interior de la clula.
Cuando en una superficie acuosa se vierte una pequea
cantidad de lpidos que tienen porciones hidroflicas e
hidrofbicas, las molculas se distribuyen uniformemente
formando una sola capa, en la cual las porciones
hidroflicas se ponen en contacto con el agua y las
porciones hidrofbicas se alejan de la superficie acuosa.
Basndose en este principio, en 1917, I Langmuir utiliz
un artefacto sumamente ingenioso denominado el recipiente
de Langmuir, con la finalidad de medir el rea de superficie
mnima ocupada por una fina capa monomolecular de
lpidos y las fuerzas necesarias para comprimir todas las
molculas de lpidos en esa rea. La utilizacin del
recipiente de Langmuir sent las bases para realizar una
serie de estudios relacionados con mediciones fsicas de los
lpidos de la membrana.
E. Gortner y F. Grandel, en 1925, publicaron sus
investigaciones sobre la organizacin de los lpidos en la
estructura de la membrana celular de los eritrocitos. Los
eritrocitos, glbulos rojos o hematies han sido y son las
clulas ms estudiadas para entender la estructura
morfolgica, la composicin bioqumica y las funciones de
la membrana. Los investigadores mencionados estudiaron
los eritrocitos de varios mamferos: cobayos, perros,
conejos, cabras, carneros y humanos empleando el
recipiente de Langmuir para concluir que la membrana
celular estaba constituida por una doble capa de lpidos
proponiendo que los extremos polares (hidroflicos) de una
capa (la externa) estaban orientados hacia afuera de la
clula y los extremos polares de la capa interna se
incorporaban hacia el interior, en contacto con la
hemoglobina; los extremos no polares de las dos capas se
dirigan hacia la parte media, quedando frente a frente como
se observa en la figura Biol. Cel.
Un modelo de membrana propuesta por James F. Danielli y
Hugh Davson en el ao de 1936, consideraba que la
membrana plasmtica estaba integrada por dos capas de
lpidos y dos de protenas, las protenas cubriendo a la de
lpidos y, cada par de ellas orientada hacia el ambiente
externo de la clula y el otro par hacia el interior
citoplasmtico como se muestra en la figura Biol. Cel. En
cada ciertos trechos de la membrana celular se insertan
protenas polares que al formar poros o canales relacionan el
ambiente externo con el citoplasmtico, permitiendo el
transporte selectivo de molculas de diversa calidad, tamao
y propiedades de solubilidad y tambin el transporte de
iones de diferente carga. (22 de julio 20:40 horas).
Biol. Cel. 8. Esquema que representa los componentes de la membrana
celular segn la teora de Danielli y Davson.
UNIDAD DE MEMBRANA DE ROBERTSON.
En la dcada de 1950 se perfeccion el microscopio
electrnico; a travs de este instrumento se lograron obtener
imgenes de componentes de la clula con un gran poder de
resolucin. A fines de esa dcada J. D. Robertson,
utilizando como fijador el tetraxido de osmio, demostr
que la imagen de la membrana plasmtica exhiba un
aspecto trilaminar caracterstico consistente en dos capas
electronlcidas u osmioflicas, una externa y una interna,
cada una de ellas constituidas por porciones polares de las
protenas y los lpidos y una capa central electronlcida u
osmiofbica integrada por los extremos no polares de los
lpidos. Las capas oscuras o electrondensas medan de 20
a 25 aproximadamente y la capa clara o electronlcida de
25 a 30 , mostrando la membrana celular, en conjunto,
un grosor de 65 a 85 . Dimensiones que coincidan con
los resultados obtenidos mediante los estudios qumicos.
Este modelo de la membrana celular se mantuvo vigente
hasta finales de la dcada del 1960. La estructura
trilaminar de la membrana plasmtica fue considerada por
Robertson y colaboradores tambin como la que constituye
las membranas de los otros componentes citoplasmticos de
la clula tales como la cubierta nuclear y las cisternas
membranales del retculo endoplsmico rugoso.
MODELO DEL MOSAICO FLUIDO DE LA
ESTRUCTURA MEMBRANAL.
En la dcada del 1970, S. Jonathan Singer y Garth
Nicholson propusieron el modelo del mosaico fluido para
explicar la estructura molecular de la membrana celular.
Este modelo considera que la membrana posee una doble
capa de lpidos en las cuales se encuentran unidas o
incrustadas molculas protenicas. Existen protenas que se
unen a las porciones polares de los lpidos, por lo tanto su
unin no es tan intensa, a estas protenas se les denominan
extrnsecas o perifricas; en otras protenas atraviesan, de
lado a lado, las dos capas de lpidos, en tanto que otras
molculas de protenas se insertan en una de las capas de
lpidos (fig. Biol. Cel. ), a estos dos tipos de protenas se les
conocen como intrnsecas o integrales.
En la superficie externa de la membrana celular, tanto en las
molculas lipdicas como en las protenicas se enlazan
molculas de carbohidratos constituyendo complejos de
glicolpidos y glicoprotenas, respectivamente.
fosfatidil inositol, fosfatidil colina, y esfingomielina.
Intercaladas entre los fosfolpidos se sitan molculas de
colesterol.
Figura Biol. Cel. 10. Disposicin espacial de las molculas de fosfolpidos
en la membrana celular. Estructura molecular de la fosfatidilcolina.(fig.
4.2 de Smith y Wood)
Biol. Cel. 9. Estructura molecular de la membrana celular segn la teora
del mosaico fluido.
El trmino de mosaico fluido, nos permite entender que las
bicapas de lpidos y protenas no son estructuras estticas
sino sumamente dinmicas, en las cuales sus componentes
son mviles, con capacidad para trasladarse de una regin a
otra de la membrana, de girar, de reunirse e interaccionar de
forma transitoria o semipermanente con otras molculas,
seales o clulas de diverso tipo.
ESTRUCTURA MOLECULAR DE LA MEMBRANA
CELULAR.
De acuerdo a lo expresado en prrafos anteriores, la
membrana celular o plasmalema est constituida por una
bicapa de lpidos (fosfolpidos) a la que se integran
protenas perifricas e integrales y molculas de
carbohidratos.
A) Fosfolpidos de las membranas.
Cada molcula de fosfolpidos (anfiptica) est compuesta
por una cabeza polar localizada en la superficie de la
membrana y dos colas largas de cidos grasos
(aciloadiposas) no polares orientadas hacia el interior del
plasmalema. Las colas no polares se disponen unas frente a
otras dentro de la membrana y se mantienen juntas por
uniones dbiles no covalentes.
Las cabezas polares estn formadas por glicerol, a ste se le
enlazan grupos nitrogenados de carga positiva mediante
grupos fosfato de carga negativa. Al glicerol se unen las dos
colas de cidos grasos mediante enlaces covalentes. De las
dos colas de cidos grasos una de ellas generalmente est
saturada y la otra no. La saturacin o insaturacin de las
colas produce una mayor o menor flexibilidad o fluidez de
la membrana. Los fosfolpidos de la membrana son
generalmente fosfatidil etanolamina, fosfatidil serina,
En las capas de fosfolpidos que constan exclusivamente de
cidos grasos saturados, las cadenas laterales estn alineadas
unas tras otras en una disposicin ordenada y cristalina
dando como resultado una estructura bastante rgida o poco
fluida, en cambio, en las capas de fosfolpidos donde existen
entremezcladas cadenas de fosfolpidos saturados e
insaturados, la fluidez y flexibilidad de la membrana son
ms evidentes. Los enlaces de tomos de carbonos, en las
cadenas insaturadas son dobles, esto le proporciona a la
cadena cierta curvatura ocasionando deformaciones
estructurales que impiden la formacin de una estructura
rgida o cristalina. La presencia de molculas de colesterol
entre las molculas de fosfolpidos le confieren una menor
flexibilidad o fluidez a la membrana (fig. Biol. Cel. ).
a)
b)
Figura Cel. Biol. 11. Disposicin de los componentes lipdicos del
plasmalema en a) membrana poco fluida, b) membrana fluida.(fig. 15.15 de
Biol. Cel. Sheeler)
La fluidez que le confieren la presencia de un nmero
mayor de molculas de fosfolpidos insaturados y la menor
cantidad de molculas de colesterol a la membrana celular,
es de tal grado que una sola molcula de fosfolpidos puede
moverse, en sentido lateral, varias micras en uno dos
segundos o trasladarse de un extremo a otro de la clulas en
pocos instantes; a este movimiento se le denomina difusin
lateral.
Existe otro movimiento de los lpidos denominado flip flop,
este movimiento es mucho ms lento, pues consiste en el
traslado de molculas de fosfolpidos de la capa interna a la
externa o viceversa, se realiza mediante un giro de 180
grados. Puede realizarse en horas, das o semanas. El
movimiento de flip flop se realizar con rapidez cuando en
el organelo denominado retculo endoplsmico liso se
sinteticen lpidos integrantes de membrana. En este caso la
actividad de enzimas llamadas flipasas colaboran
activamente en desplazar molculas de fosfolpidos desde
una mitad de la membrana a la otra mitad.
Gracias a la flexibilidad de la bicapa de fosfolpidos, las
membranas celulares son fcilmente deformables y
flexibles, caracterstica que le permite a la clula cambiar de
forma (leucocitos granulocitos, eritrocitos, amibas) para
adecuarse a ciertos ambientes, modificarse durante la
divisin celular o cuando efecta
movimientos de
locomocin; facilita la fusin o el desdoblamiento de las
membranas, por ejemplo en la fecundacin del ovocito con
el espermatozoide, o en la formacin de seudpodos o
filopodios durante la fagocitosis o la formacin de vesculas
de secrecin y su posterior fusin con el plasmalema para
exocitar el producto sintetizado.
ASIMETRA DE LOS LPIDOS EN LA MEMBRANA
CELULAR:
Los diferentes tipos de fosfolpidos de la membrana celular
no se encuentran distribuidos equitativamente en las dos
capas de fosfolpidos. La distribucin de los lpidos en la
membrana del eritrocito es asimtrica tal como se observa
en la siguiente tabla:
Fosfolipidos de
membrana
Esfingomielina
Fosfatidil colina
Fosfatidil
etanolamina
Fosfatidil serina
Lpidos de la
capa externa (%)
20
23
6
Lpidos de la
capa interna (%)
6
9
25
10
Las molculas de colesterol, en cambio, tienen una
distribucin ms o menos equitativa en ambas capas.
B) Protenas de las membranas.
Protenas integrales o intrnsecas. Son protenas
generalmente helicoidales que atraviesan totalmente la
bicapa de fosfolpidos; esto da como resultado que posean
regiones hidroflicas e hidrofbicas. Aquellas porciones de
la protena inserta en la capa de fosfolpidos tienen
abundantes aminocidos con cadenas laterales hidrofbicas.
Las porciones que se proyectan hacia el exterior de la doble
capa de lpidos poseen aminocidos hidroflicos. Aquellas
que se ponen en contacto con el microambiente extracelular
se asocian con cadenas de carbohidratos (glicoprotenas). En
muchos casos las protenas integrales constituyen canales
acuosos que atraviesan la membrana y facilitan el transporte
de sustancias hidrosolubles como iones, hormonas,
partculas de bajo peso molecular, agua, vitaminas, etc. Las
protenas integrales suelen desplazarse lateralmente en el
interior de la membrana. El trmino de mosaico fluido
explica tambin esta movilidad lateral de las protenas.
B)
A)
B)
Figura Biol. Cel. 12. Disposicin espacial de las molculas protenicas
integrales en la membrana celular. A) protenas multipasos y B) protenas
de de un solo paso o paso nico.
Muchas de ellas poseen ms de un segmento dentro de la
membrana. Las cadenas de estas protenas integrales son
bastantes largas y plegadas, su disposicin dentro de la
membrana representa varios pasos a travs de ella, por esta
caracterstica se les denominan protenas multipasos o
politpicas. Los dominios citoplasmticos y extracelulares
de estas protenas poseen receptores especficos para
molculas de sealamiento particulares. En otros casos la
molcula protenica, no tan larga, slo posee un segmento
dentro de la bicapa de fosfolpidos. Se les conoce como
protenas integrales de un solo paso o bitpicas. Sus
extremos estn expuestos a ambos lados de la membrana.
Son protenas que actan como receptores que enlazan
diferentes ligandos o primeros mensajeros, en su superficie
externa por ejemplo, un factor de crecimiento, un antgeno o
una hormona peptdica, con la finalidad de transmitir un
mensaje mediante su extremo citoplasmtico.
Figura Biol. Cel. 13. Estructura molecular de los diferentes tipos de
protenas integrales y perifricas de la membrana celular.
Protenas extrnsecas o perifricas. Son protenas unidas
dbilmente a las superficies externa (extracelular) e interna
(citoplasmtica) de la bicapa de fosfolpidos, mediante
enlaces electrostticos. Estn constituidas por cadenas de
aminocidos hidroflicos que interactuan con el agua que las
rodea y con las cabezas polares de los fosfolpidos. Las
externas se asocian a cadenas de carbohidratos.
Las protenas perifricas ms estudiadas son aquellas que
estn en relacin con el citoplasma de la clula. Una de
ellas, la espectrina, forma una especie de red fibrilar
tridimensional que acta como un soporte o esqueleto al
suministrar un apoyo mecnico flexible a la membrana
cuando ocurren cambios rpidos en la morfologa celular o
intervienen como enlaces para conectarse a una red fibrilar
tridimensional protenica citoplasmtica que integran el
denominado citoesqueleto o se vinculan con el sistema de
segundos mensajeros.
Figura Biol. Cel. 14. Disposicin espacial de las protenas integrales y
perifricas del eritrocito
Asimetra de las protenas membranales. Las regiones
externas e internas de la membrana celular poseen diferentes
tipos y cantidades de protenas perifricas o integrales. La
membrana del eritrocito posee mayor cantidad de protenas
en su capa interna y un menor nmero de ellas existen en la
capa externa.
Uno de los procedimientos que facilit la observacin de
esta asimetra de las protenas en las dos capas fue el de
criofracturacin de membranas + sublimacin de metales
y posteriormente obtener imgenes de ellas mediante el
microscopio electrnico de transmisin.
Figura Biol. Cel. 15. Imagen de una membrana criofracturada mostrando
la distribucin de las protenas integrales.
Para obtener las imgenes, las membranas celulares se
congelan rpidamente con nitrgeno lquido, a continuacin
se fracturan con una cuchilla especial de ultramicrotomo
siguiendo planos especficos. Cuando el plano de fractura
coincide con la porcin electronlcida de la membrana, sta
se separa en dos a lo largo de la bicapa lipdica. Una vez
fracturada la membrana se deposita, sobre las superficies
expuestas, metales pesados como el platino, encima de esta
cubierta metlica se esparcen tomos de carbono,
obtenindose rplicas en relieve, para ser observadas con el
microscopio electrnico. Las superficies expuestas se
denominan: superficie E, aquella relacionada con la mitad
externa y P la superficie que est en contacto con el
protoplasma (mitad interna). Las imgenes obtenidas
muestran una superficie llena de elevaciones irregulares de
diverso tamao intercaladas con pequeas depresiones o
crteres (fig. Biol. Cel. ) que permiten analizar la
heterogeneidad de la distribucin de las protenas en ambas
capas de la membrana.
C) Carbohidratos de la membrana celular.
Uno de los componentes de la membrana celular es la
presencia de carbohidratos. Suelen ser de cadenas cortas o a
veces ramificadas. Los principales azcares se derivan de la
glucosa. Todos ellos se forman como disacridos por unin
de seis azcares: D-galactosa, D-manosa, L-fucosa, cido
N.acetlneuramnico o cido silico, N-acetil-Dglucosamina y N-acetil-D-galactosamina, predominan en
ellos los grupos sulfatos y carboxilos.
Los carbohidratos se enlazan de forma covalente con los
fosfolpidos (glucofosfolpidos) y con las protenas
(glucoprotenas) de la superficie externa de la membrana.
Constituyen el glucocalix de las clulas; estn presentes en
un 2 y 10% del total del peso de la membrana. No existen
cadenas de azucares ligados a la superficie citoplasmtica de
la membrana celular. Esta disposicin estructural de los
carbohidratos afianza con mayor fuerza la denominada
asimetra de la membrana celular en sus componentes
qumicos.
Cuando se observan imgenes electrnicas de superficies
celulares, el glucoclix se muestra como una capa vellosa o
algodonosa localizada encima de la lmina electrondensa
superficial (fig. Biol. Cel. ).
El cido silico es el azcar terminal de los oligosacridos
del glucoclix y le confiere carga negativa a las cadenas de
carbohidratos. Esta caracterstica, unida a la presencia de
grupos sulfatos y carboxilos propicia que reaccionen
positivamente con el cido peridico + el reactivo de
Schiff, colorendose de rojo magenta; con el azul alciano, se
tien de color azul celeste y de color rojo con el rojo de
rutenio. Las clulas intestinales de absorcin y las clulas
del tbulo contorneado proximal del parnquima renal,
poseen abundantes microvellosidades en su superficie
apical, las cuales estn cubiertas con una cantidad
considerable de glucoclix.
Figura Biol. Cel. 16. Glucocalix imagen electrnica del glucocalix
presente en las microvellosidades de clulas intestinales.
El glucoclix desempea varias funciones: Una de las ms
importantes es la de proteger a las clulas de la interaccin
de protenas inapropiadas o de lesiones qumicas o fsicas.
Los glucolpidos de la superficie de los eritrocitos
determinan si el grupo sanguneo de una persona es A, B,
AB O. La presencia de los oligosacridos determina la
especificidad celular, en consecuencia intervienen en el
reconocimiento y adherencia de clulas con clulas para
mantener la cohesin en los tejidos o en las reacciones
antignicas de las membranas celulares, desencadenando
una respuesta inmunolgica.
Los azcares membranales tienen una gran afinidad por las
lectinas, un tipo especial de protenas vegetales que se
encuentran principalmente en las leguminosas. Se combinan
entre s de una manera semejante a la unin de una enzima
con su substrato o de un anticuerpo con su antgeno. Los
diferentes tipos de carbohidratos interaccionan de forma
especfica con un determinado tipo de lectinas; esta
especificidad se utiliza para localizar y mapear la
distribucin de los azcares en la superficie celular. La
interaccin entre lectinas y carbohidratos especficos, suele
emplearse para diferenciar las membranas de clulas
normales de las clulas malignas o cancerosas que derivan
de aquellas. Esto se debe a que las membranas de las clulas
cancerosas son ms fluidas y facilitan que las lectinas
puedan agrupar con ms facilidad las glucoprotenas de las
clulas metaplsicas y reunirlas o aglutinarlas en grupos de
clulas y propiciando la precipitacin de ese grupo celular.
Transporte de sustancias a travs de la membrana
celular.
La membrana celular es una barrera de permeabilidad
selectiva. A travs de ella se transportan una serie de
sustancias desde el microambiente extracelular hacia el
interior citoplasmtico y viceversa.
La clula requiere del transporte a travs del plasmalema
para poder realizar sus procesos metablicos; gracias a ellos,
los organismos unicelulares y pluricelulares
pueden
mantener un equilibrio u homeostasis, es decir una
regulacin equilibrada del metabolismo (anabolismo y
catabolismo) para mantener las condiciones adecuadas de la
vida.
Para mantener este equilibrio las clulas necesitan del
suministro o aporte de sustancias del medio que las rodea:
agua, gases, azcares, lpidos, aminocidos, vitaminas,
hormonas y sales. As mismo, la clula sintetiza y secreta
sustancias y requiere excretar desechos, productos del
catabolismo celular. Estas funciones estn reguladas y
coordinadas por la capacidad que tiene el plasmalema de
seleccionar dentro de una gran cantidad de molculas y
iones slo aquellos que necesita la clula para mantener sus
funciones vitales tales como el crecimiento, la reproduccin,
la sntesis de sustancias, la respiracin, etc.
Tipos de transporte celular. El traslado de sustancias
del medio extracelular al interior de la clula y viceversa se
lleva a cabo por medio de los mecanismos siguientes:
Transporte pasivo. Este transporte se sustenta en la
capacidad del movimiento o energa cintica de las
partculas que atraviesan la membrana, por lo cual la clula
no tiene que utilizar energa para que se produzca este
proceso.
Transporte activo. En este caso la clula debe invertir
energa para transportar molculas y iones que no
atravesaron la membrana mediante el transporte pasivo.
Endocitosis. Es el proceso mediante el cual la clula ingiere
sustancias lquidas, macromolculas y material en forma de
pequeas partculas provenientes del ambiente extracelular
y los transporta al citoplasmtico. La endocitosis es de dos
tipos:
a) Fagocitosis,
cuando
la
clula
incorpora
macromolculas y partculas slidas como bacterias,
eritrocitos, stas deben medir un tamao mayor de 250
nanmetros de dimetro. etc.
b) Pinocitosis, se produce cuando la clula ingiere o
bebe gotitas de lquidos, generalmente de un dimetro
menor a 150 nm.
c) Exocitosis. Es el proceso mediante el cual la clula
libera hacia el exterior sustancias que sintetiza
(secrecin) o desechos que debe eliminar (excrecin).
FACTORES QUE INTERVIENEN PARA QUE EL
TRANSPORTE CELULAR SE LLEVE A CABO:
El tamao de las molculas. Molculas pequeas como
las sales y algunas molculas orgnicas atraviesan el
plasmalema con ms facilidad que molculas de gran
tamao como las protenas.
La carga elctrica de las partculas. Las molculas sin
carga elctrica atraviesan ms fcilmente la membrana
que aquellas que la poseen. An con carga elctrica las
partculas con menor carga, como las monovalentes (Cl -)
atraviesan el plasmalema con mayor facilidad que
aquellas de mayor carga, pero del mismo signo, como
por ejemplo los divalentes o trivalentes, por ejemplo
( PO43_ ).
El nmero de molculas de agua. Las partculas que
posean una mayor cantidad de agua ligadas a su
superficie pasarn con ms facilidad la membrana.
La solubilidad de las partculas en lpidos. Cuanto
mejor se disuelvan las partculas en lpidos atravesarn
la membrana con mayor rapidez.
Transporte pasivo:
El transporte pasivo se realiza sin necesidad que la clula
utilice energa, Existen varias formas de este transporte:
Difusin simple.
Se define la difusin como el movimiento espontneo y
aleatorio de molculas o partculas de una zona de mayor
concentracin a otra de menor concentracin, es decir a
favor de un gradiente de concentracin hasta que las
concentraciones de la sustancia se equiparen en ambas
zonas
(que alcancen un equilibrio dinmico). De esta
manera atraviesan la membrana sustancias apolares como
el oxgeno, el benceno, el xilol o el nitrgeno y molculas
polares pero sin carga como el agua.
Figura Biol. Cel. 17. Esquema que representa el transporte pasivo por
difusin simple.
Difusin de iones a travs de la membrana. Los lpidos que
integran la bicapa hidrfoba de las membranas es bastante
permeable a sustancias con carga elctrica incluyendo iones
pequeos como Na+, K+, Ca2+ y Cl -. A este tipo de
transporte de iones se les denomina a travs de canales
inicos. En la actualidad se han identificado un nmero
bastante grande de canales inicos, cada uno formado por
protenas integrales que rodean un poro acuoso. Estos
canales son sumamente especficos y nicamente dejan
pasar un solo tipo de iones. El transporte de iones siempre lo
hacen a favor de un gradiente de concentracin o cuesta
abajo es decir de un estado de mayor energa hacia otro de
menor energa. Los canales inicos son bidireccionales
pues permiten el transporte de iones en ambas direcciones,
el flujo de los iones depende del gradiente electroqumico.
La importancia de los canales inicos radica en que
desempean mltiples funciones celulares relacionadas con
la generacin y propagacin de impulsos nerviosos,
contraccin muscular, secrecin de sustancias hacia el
espacio extracelular, y la regulacin del volumen celular.
La velocidad de difusin de las partculas depende
a) del estado de agregacin de sus tomos (slido,
lquido o gaseoso), la difusin es ms rpida en gases
(O2, CO2, Nitrgeno);
b) su masa, a mayor masa la difusin es ms lenta,
c) la concentracin de las partculas, a mayor
concentracin de partculas en el medio, mayor
velocidad de difusin y
d) la temperatura ambiente, conforme aumenta la
temperatura la energa cintica de las partculas
aumenta, en consecuencia tambin su velocidad de
difusin.
Figura Biol. Cel. 18. Difusin a travs de canales inicos.
Los canales inicos protenicos son semejantes entre s, su
conformacin qumica est constituida por una secuencia de
aminocidos similares en todos ellos, tanto en clulas
bacterianas, vegetales y animales, indicndonos que
provienen de un ancestro comn, a pesar que cada canal
inico es especfico para el transporte de determinados
iones.
La mayora de los canales inicos suelen poseer una
conformacin cerrada o abierta. Actan como compuertas
de la clula que pueden abrirse o cerrarse de acuerdo a los
siguientes tipos de canales:
Canales operados por compuertas de voltaje. Su
conformacin depende de la carga elctrica que exista en
ambas superficies de la membrana. Los canales se abren
y dejan pasar iones como el Na + para que se produzca la
despolarizacin de la membrana en la transmisin de los
impulsos nerviosos; en estos casos la posicin del canal
abierto es inestable y despus del pasaje del ion, el canal
se cierra y no se abre hasta que transcurran algunos
milisegundos (periodo refractario).
Canales operados por compuertas de sustancias
qumicas o ligandos. La conformacin de ellos depende
del enlace qumico que se establece con una sustancia en
particular denominado ligando o molcula seal. El
canal se abre y permanece abierto hasta que la molcula
qumica o ligando se disocia de la protena del canal, por
ejemplo los neurotransmisores, stos generalmente
llegan a la membrana postsinptica de las uniones
neuronales (axn-dendrita o axn-clula efectora) o
pueden ser nucletidos como el cAMP (el monofosfato
de adenosina cclico) en las neuronas olfatorias y el
cGMP (el monofosfato de guanosina cclico) que acta
en los bastones de la retina.
Canales con compuerta mecnica. En este tipo de
canal se necesita una maniobra fsica o mecnica real
para abrir la compuesta. Este mecanismo acta en las
clulas pilosas del odo interno (vestibulares o
cocleares), localizadas en el epitelio sensorial de los
conductos vestibulares o del rgano de Corti,
respectivamente. Poseen en sus porciones apicales una
serie de microvellosidades (estereocilios) que se doblan
cuando sobre ellas vibra una sustancia gelatinosa (la
cpula o la membrana tectoria), al doblarse la membrana
celular abre los canales y los iones penetran al
citoplasma produciendo despolarizacin y la transmisin
del impulso nervioso, ste llega al encfalo y se traduce
en sensaciones de movimiento, orientacin o sonidos.
Canales con compuertas de protenas G. Ciertos
canales como aquellos que dejan pasar la acetilcolina
para la contraccin del msculo estriado cardiaco
necesitan la interaccin de una molcula receptora y una
protena G. Las protenas G, son enzimas trimricas
compuestas por de una subunidad alfa, grande y dos
subunidades pequeas: beta y gamma y se relacionan a
receptores. Existen varios tipos de protenas G,
estimuladoras (Gs) e inhibitorias (Gi ). Las protenas G
son activadas por la sustancia aceptada en el receptor
propiciando la modulacin del canal protenico para que
ste pueda abrirse o cerrarse.
Canales sin compuerta. Un ejemplo de estos canales
es aquel que facilita el escape de iones potasio (K + );
indispensable en la creacin de un potencial elctrico
(voltaje) entre las dos superficies de la membrana
celular. Al carecer de compuerta, la clula no puede
controlar el pasaje de iones de K + de esta manera el
trnsito de los iones de potasio refleja la concentracin
de este ion en ambos lados de la membrana.
Difusin facilitada.
Se le denomina as a otra forma de difusin que se produce
en la clula a favor de un gradiente de concentracin, es
decir de una regin de mayor concentracin a otra de menor
concentracin empleando para tal fin molculas protenicas
transportadoras o acarreadoras localizadas en la
membrana. La presencia de estas protenas ayudan a que el
transporte sea ms rpido que la difusin simple. Las
protenas transportadoras slo pueden enlazarse a un soluto
cada vez, en un lado de la membrana en la que un cambio de
conformacin acta como un mecanismo para transportar la
sustancia a travs de la membrana. El transporte de las
sustancias puede modularse de acuerdo a los requerimientos
de la clula Un ejemplo de este transporte se realiza en la
entrada de la glucosa hacia el interior de las clulas (fig.
Biol. Cel. ). Este tipo de protena transportadora se
denomina uniporte, es decir el traslado de sustancias se
hace en una sola direccin del exterior hacia el citoplasma.
Figura Biol. Cel. 19. Transporte mediado por protenas acarreadoras.
Un tipo especial de canal existente en ciertas clulas son los
canales de agua por ejemplo, en la superficie apical de
las clulas de los tbulos contorneados proximales del
rin; all existen unas protenas transportadoras
denominadas acuaporinas que actan como un sistema
especializado y sumamente eficaz en el transporte de agua.
smosis. Un caso especial de difusin es la smosis,
proceso que consiste en el paso de un solvente a travs de
una membrana semipermeable desde una regin de mayor
concentracin de agua (menor concentracin de soluto) a
otra de menor concentracin de agua (mayor concentracin
de soluto). El agua es la sustancia que penetra o sale de la
clula mediante este proceso.
La presin que ejerce el agua en el interior de la clula se
denomina presin osmtica y es aquella que permite o no la
salida del agua para poder igualar la concentracin de agua
en el exterior de la clula.
Cuando la concentracin de agua y soluto es similar en el
exterior como en el interior de la clula, se dice que el
liquido extracelular es isotnico o isosmtico con respecto
a la clula, condicin ideal para que en ella se produzcan de
manera normal todas las funciones vitales; por ejemplo,
cuando eritrocitos se suspenden en una solucin de cloruro
de sodio al 0.9%, en ellos no se produce ninguna
modificacin porque esta concentracin de Cl 2Na es la que
existe en el plasma sanguneo. En cambio, si en el medio
extracelular existe una mayor concentracin de soluto y una
menor de agua, el medio sera
hipertnico o
hiperosmtico con respecto a la clula, por lo tanto sta
pierde agua para igualar las dos concentraciones lo que
produce prdida de volumen y la clula se encoge o arruga
(crenacin), por ejemplo cuando eritrocitos se suspenden en
una solucin de cloruro de sodio al 1.3%. Si sucede el caso
contrario, es decir que en el medio externo exista una mayor
concentracin de agua y menor cantidad de soluto, la clula
se encuentra rodeada de un medio hipotnico o
hipoosmtico con respecto a ella, esto ocasionara que el
agua penetre al interior de la clula para equilibrar
adecuadamente la disolucin del soluto en el citoplasma,
produciendo el aumento del volumen celular por ganancia
de agua; si no se igualan las concentraciones existe la
posibilidad que la clula contine aumentando de volumen
hasta que la membrana se rompa o se debilite de tal forma
que ahora permita la salida del soluto para que se igualen las
concentraciones del mismo en los mbitos: extra e
intracelular. Esto sucede cuando los eritrocitos se colocan en
una solucin de cloruro de sodio al 0.6%; en este caso los
eritrocitos se hacen esfricos, la membrana celular se
debilita y la hemoglobina sale al medio exterior (hemlisis).
Transporte activo.
La concentracin de iones en el mbito citoplasmtico de la
clula se conserva elevada gracias a un sistema de bombeo o
transporte activo generado a travs del plasmalema en
contra de un gradiente de concentracin, esto significa que
se trasladan iones y molculas de una regin de menor
concentracin a una de mayor concentracin. Ciertas
molculas con carga elctrica difcilmente atravesaran la
membrana en esas condiciones mediante difusin simple o
facilitada. Los aminocidos, el potasio ( K +), el magnesio
(Mg2+) y el Na+ son transportados a travs de la membrana
desde regiones de baja concentracin a otras de
concentracin elevada utilizando energa, es decir mediante
transporte activo.
Se denomina transporte activo al movimiento de iones
o molculas, a travs de la membrana, en contra de un
gradiente de concentracin debindose invertir para
tal fin, parte de la energa obtenida en el metabolismo.
Un ejemplo de este tipo de transporte es el mantener la
concentracin de potasio y de sodio dentro del citoplasma
celular. En la especie humana y en los mamferos la
concentracin de potasio dentro de las clulas es 50 100
veces mayor que en el plasma sanguneo y el sodio est en
una concentracin muy alta fuera de la clula. Para
mantener las concentraciones diferenciales de Na+ y K+ la
clula debe gastar adenosina trifosfato (ATP) para impulsar
una protena transportadora antiporte acoplada conocida
como bomba de Na+ - K+. Esta bomba transporta iones de
K+ hacia el interior de la clula y iones de Na + hacia el
exterior, en ambos casos contra un gradiente de
concentracin. La membrana de las clulas animales posee
una gran cantidad de bombas de sodio y potasio pues
mantener sus concentraciones equilibradas. El equilibrio de
estos iones es esencial e indispensable para la supervivencia
y funcionamiento normal de la clula. El tipo de transporte
por el cual a travs de un solo canal se trasladan en sentidos
opuestos dos tipos distintos de iones Na+ - K+ se le
denomina antiporte.
Figura Biol. Cel. 20. Eritrocitos suspendidos en a) solucin isotnica, b)
solucin hipertnica y c) solucin hipotnica.
La smosis es un factor importante para las funciones del
organismo. Por ejemplo, el aparato digestivo secreta una
gran cantidad de lquidos: jugos gstricos, intestinales,
pancreticos y bilis, y alberga lquidos que llegan junto con
los alimentos, estos lquidos deben reabsorberse
osmticamente en el epitelio intestinal. Si este proceso de
reabsorcin no ocurre, se produce una diarrea sumamente
profusa generndose una rpida deshidratacin
que
atentara con la vida de una persona o animal.
Figura Biol.Cel. 21. Esquema que representa el transporte activo.
La protena integral de transporte que acta como de bomba
de Na+ y K+ tiene dos sitios de enlace para el K + en su lado
externo y tres lugares en la superficie citoplasmtica para el
Na+, en consecuencia cuando se acarrean dos iones de K +
hacia el interior de la clula, se transportan tres iones de Na +
hacia el lado de afuera.
La enzima ATPasa de Na+ - K+ interviene en la bomba de
Na+ - K+. Cuando se ligan tres iones de Na+ en la superficie
citoslica de la bomba, el ATP se hidroliza en difosfato de
adenosina (ADP) y el ion fosfato liberado se utiliza para
fosforilar la ATPasa lo que genera la modificacin de la
configuracin de la bomba con la consiguiente transferencia
de iones Na+ fuera de la clula. La unin de dos iones de K +
en la superficie externa de la membrana causa
defosforilizacin de la ATPasa con el retorno subsiguiente
de la protena transportadora a su configuracin anterior
propiciando la transferencia de iones K + hacia el interior de
la clula.
El funcionamiento constante de la bomba sodio-potasio
reduce la concentracin inica citoplasmtica y tiene como
efecto reducir la presin smotica intracelular. Si esta
reduccin osmtica no se llegara a producir entonces
entrara agua a la clula en grandes cantidades produciendo
un exceso de turgencia en la clula y su consecuente
estallamiento.
COMPONENTES DEL CITOPLASMA.
Los procesos metablicos en la clula animal, es decir, los
procesos de intercambio de materia y energa con el
ambiente que las rodea estn integrados por mecanismos
muy complejos, resultantes de la sincronizacin y el
acoplamiento de reacciones qumicas en las que intervienen
una serie de molculas y compuestos orgnicos e
inorgnicos, generando como consecuencia los procesos
vitales de la clula.
Los elementos qumicos ms abundantes de las clulas son
carbono, hidrgeno, oxgeno y nitrgeno. Estos elementos
constituyen casi el 96% del peso total de la clula..
En las clulas se encuentran dos tipos de compuestos:
inorgnicos y orgnicos.
a) Los compuestos inorgnicos son aquellos que no poseen
enlaces carbono-carbono. Los ms comunes son el agua; los
gases disueltos como el oxgeno (O 2) y el dixido de
carbono (CO2); sales minerales con elementos
constituyentes de fierro, fsforo, calcio, magnesio, potasio,
sodio, cloro, cobre, zinc, y manganeso.
b) Los compuestos orgnicos estn constituidos por
cadenas de tomos de carbono enlazados con uno o ms
elementos como el oxgeno, hidrgeno, nitrgeno, azufre y
fsforo. Los principales compuestos orgnicos de las clulas
son carbohidratos, protenas, lpidos, cidos nucleicos y
vitaminas.
Agua.
El hidrgeno y el oxgeno se combinan entre s para formar
el compuesto inorgnico ms abundante en las clulas y, por
consiguiente, en todos los seres vivos. En los mamferos
(humanos inclusive) casi el 80% del cuerpo es agua. Por lo
expuesto, podemos afirmar que el componente principal del
citoplasma es el agua. Todos los procesos vitales de la clula
se producen en presencia de agua Sin agua no existe la
vida.
PROPIEDADES DEL AGUA.
Est considerada como el solvente de la vida, disuelve
una gran cantidad de sustancias. Esto se debe a su estructura
polar; en ella los electrones compartidos del hidrogeno son
atrados hacia un ncleo del oxgeno, presentando una
polaridad negativa del lado del oxgeno y polaridad positiva
del lado del hidrgeno, con lo cual las molculas de agua
pueden orientarse hacia iones positivos y negativos. El
carcter excepcional de solvente le permite al agua ser el
vehculo por medio del cual las sustancias disueltas en ella,
pueden atravesar la membrana de la clula, hacia su interior,
para nutrirla, o hacia el exterior para secretar sustancias o
eliminar sus desechos.
El agua es un regulador de la temperatura. Es un
compuesto muy estable debido a su alta capacidad
calorfica, es decir, absorbe una gran cantidad de calor antes
de liberarlo en su entorno, esta propiedad permite que el
agua mantenga constante la temperatura de la clula donde
muchos organismos puedan continuar viviendo sin
congelarse, evitando la prdida de calor.
Permanece en estado lquido en un amplio rango de
temperaturas, desde 0o (temperatura en la adquiere el estado
slido: hielo) hasta 100o (temperatura en la que hierve y se
transforma en gas). Hecho importante para los seres vivos
pues para elevar la temperatura del agua al punto de
ebullicin y vaporizacin se requiere utilizar mucha energa.
Si esto no ocurriera as, probablemente el agua se
encontrara en forma de vapor a la temperatura ambiente,
estado que ocasionara la muerte de las clulas.
Es un excelente lubricante en los organismos. Forma parte
de todos los lquidos tisulares, evita la friccin entre
superficies tisulares que tienen contacto con otras como en
el caso de las articulaciones.
Participa en la gran mayora de las reacciones
bioqumicas. Algunas de ellas son la respiracin celular y
la fotosntesis.
COMPUESTOS ORGNICOS MS IMPORTANTES
DE LAS CLULAS:
Carbohidratos: Tambin se les conoce con el nombre de
sacridos, azcares o glcidos. Constituyen una reserva
energtica de las clulas. Su estructura qumica esta
compuesta por el Carbono (C), el Hidrgeno (H) y el
oxgeno (O).
En la clula animal, la glucosa (monosacrido) es la
principal fuente de energa. Cuando se polimeriza forma un
polisacrido, denominado glucgeno que se almacena como
reserva de energa en varios tipos celulares: las fibras
musculares esquelticas o del corazn y en los hepatocitos
(clulas del hgado).
La ribosa y la desoxirribosa son azcares monosacridos
que integran la molcula de los cidos nucleicos: el cido
ribonucleico y el desoxirribonucleico, respectivamente.
La lactosa es un azcar disacrido, que forma parte de la
secrecin lctea (leche) de los mamferos. Resulta de la
unin de dos monosacridos, la glucosa y la galactosa,
sintetizada en las clulas de la glndula mamaria. Tambin
es una fuente de energa.
Protenas. Son macromolculas formadas por tomos de
carbono (C), hidrgeno (H), oxgeno (O), nitrgeno (N) y
azufre (S), en ciertos casos se le unen tomos de hierro (Fe),
zinc (Zn) y cobre (Cu). Estn constituidas por molculas de
menor tamao llamadas aminocidos. Forman parte de
componentes estructurales y funcionales esenciales de la
clula: de las membranas celulares, el citoesqueleto,
sustancias que constituyen parte de secreciones e integran
molculas hormonales y anticuerpos.
Los aminocidos son molculas que poseen un grupo cido
denominado carboxilo (COOH), y un grupo bsico, llamado
amino (NH2). Cuando los aminocidos se unen para formar
protenas, se enlazan el grupo amino de uno con el grupo
carboxilo del otro, establecindose un enlace denominado
peptdico con la liberacin de una molcula de agua (H2 O).
La unin de aminocidos da como resultado la formacin de
molculas ms complejas as tenemos que cuando se unen:
Aminocido + aminocido = dipptido
Aminocido + dipptido = oligopptido
Ms de 10 aminocidos = polipptido.
Polipptidos + polipptidos = protena.
Estructura espacial de las protenas:
De acuerdo a la organizacin de los aminocidos y su
acomodacin en el espacio, las protenas se clasifican en:
Estructura primaria. Esta dada por la secuencia de
aminocidos unidos por enlaces peptdicos. Tiene un
aspecto lineal.
Estructura secundaria. Mediante enlaces de puentes de
hidrgeno se establecen uniones con los aminocidos de la
cadena peptdica, este arreglo le confiere a la protena
patrones de plegamientos en forma de hlices (de espiral)
con giros a la derecha o a la izquierda.
Estructura terciaria. Es la disposicin de los pptidos que
le proporciona la estructura tridimensional a la protena.
Est dada por el plegamiento global de la cadena
polipptidica.
Estructura cuaternaria. Es proporcionada por la unin de
dos o ms cadenas polipptidicas individuales que se unen
para funcionar como una sola unidad. En ciertos casos la
unin de estas cadenas es indispensable para el
funcionamiento adecuado de la protena.
La forma o el plegamiento de una protena se puede alterar
cuando en el medio intracelular o extracelular se producen
modificaciones en la temperatura, el pH (cambios en la
acidez o alcalinidad del medio) y las concentraciones de
sales. Cuando estas modificaciones ocurren entonces se
afirma que la protena se ha desnaturalizado y ocasionar
una alteracin en la forma o en el funcionamiento de la
clula que la contiene.
Clasificacin de las protenas. Se clasifican de acuerdo a
tres criterios:
Su composicin qumica. Pueden ser a) simples,
cuando estn constituidas solamente por aminocidos,
como la insulina, hormona pancretica que interviene en
el metabolismo de la glucosa y b) conjugadas, en este
caso la protena se une a un carbohidrato para integrar
una glucoprotena o se une a un lpido para constituir
una lipoprotena; o a un elemento inorgnico, ejemplo el
hierro (Fe) como en el caso de la hemoglobina.
Su forma. Se clasifican en a) globulares, por la forma
esfrica que adoptan, ejemplos, la actina, protena
contrctil de las clulas (fibras) musculares; la
hemoglobina, protena de los eritrocitos, responsable del
transporte de O2 y CO2, y b) fibrosas. Estn compuestas
por cadenas filamentosas, alargadas. Abundan en
muchas clulas como la protena queratina en las clulas
epiteliales de la piel, o forman parte de sus secreciones,
por ejemplos: la protena colgena o la elastina,
sintetizadas y secretadas por fibroblastos.
Su funcin biolgica. De acuerdo a la funcin que
realizan las protenas son:
a) estructurales, son todas aquellas protenas que
constituyen la estructura morfolgica de las clulas
como
las
protenas
membranales,
los
microfilamentos y microtbulos, la queratina, el
humor vtreo del globo ocular; las fibras colgenas
y elsticas que forman tendones y ligamentos; b)
Funcionales, promueven una variedad de
funciones celulares
b) Catalizacin y digestin; las enzimas, porque
aceleran la velocidad de las reacciones qumicas de
las clulas, por ejemplo las hidrolasas que actan
en el rompimiento de carbohidratos y protenas.
c)
Transporte. La tubulina que forma microtbulos,
encargados del transporte intracelular o la
hemoglobina que transporta el O 2 y CO2. Protenas
membranales que permiten el transporte de
sustancias del exterior de la clula al citoplasma y
viceversa.
d) Proteccin. Sntesis y secrecin de protenas en la
forma de inmunoglobulinas o anticuerpos que
intervienen en la defensa del organismo ante el
ataque de microrganismos y sustancias extraas.
e)
Contraccin. Las protenas actina y miosina,
intervienen en el movimiento contrctil de las
clulas.
f)
Receptores de estmulos y reconocimiento celular.
Protenas membranales que permiten la adhesin y
reconocimiento de clulas de estirpe semejante y la
recepcin de estmulos a travs de ligandos.
Lpidos. Tambin se les conoce con el nombre de grasas.
Los lpidos son molculas insolubles en agua pero solubles
en solventes orgnicos como el benceno, el xilol, el ter, la
acetona y el petrleo. Constituyen un grupo heterogneo de
compuestos. Estn formados por carbono (C), hidrgeno
(H), oxgeno (O), En algunos casos tambin se integran
fsforo (P) y nitrgeno (N).
Los lpidos se clasifican en:
Lpidos complejos con cidos grasos. En este grupo se
consideran a
a)
las
grasas
neutras
o
triaclglicridos
(triglicridos). Estn compuestas por dos subunidades:
el glicerol (un alcohol de tres carbonos) y los cidos
grasos (compuestos de hidrocarburos de cadena larga
que poseen un grupo carboxilo en un extremo). Se
almacenan en varios tipos celulares pero de manera
abundante en los adipocitos o clulas adiposas. En ellas
actan como reservorios de energa (un gramo de grasa
proporciona ms del doble de la energa que un gramo
de carbohidratos), de aislantes trmicos y mecnicos.
b) Fosfolpidos o fosfoglicridos. Los constituyen el
glicerol y los cidos grasos pero poseen, adems, un
tomo de fsforo (P) y un compuesto orgnico que vara
segn el tipo de fosfolpido. Son componentes
estructurales de las membranas celulares.
c) Esfingolpidos. Estn formados por un aminoalcohol
llamado esfingosina, una molcula de cido graso y una
variedad de otros compuestos. Forman parte de
membranas celulares especialmente de la vaina de
mielina de las neuronas (clulas nerviosas) sustancia
constituida por la esfingomielina que interviene
activamente en la transmisin y conduccin de los
impulsos nerviosos.
Lpidos simples sin cidos grasos. Se consideran dentro de
esta clasificacin a los
a) Esteroides. Son molculas complejas derivadas de
cuatro anillos fusionados de tomos de carbono. Un
ejemplo de este lpido es el colesterol, componente
estructural de las membranas celulares. Es un precursor de
los cidos biliares y de una serie de hormonas esteroideas
como la testosterona (hormona sexual masculina), la
progesterona y estradiol (hormonas femeninas); el cortisol
y la aldosterona (hormonas sintetizadas en la corteza
suprarrenal).
b) Prostaglandinas. Son compuestos derivados de los
cidos grasos. Participan en la contraccin muscular de las
fibras musculares lisas del aparato urogenital, por ejemplo
provocando las contracciones del tero durante el parto y,
en el metabolismo de los lpidos. Ejercen efectos
fisiolgicos y farmacolgicos de tipo hormonal.
Acidos nucleicos. Los cidos nucleicos son molculas
complejas integradas por tomos de carbono (C), hidrgeno
(H), oxgeno(O), nitrgeno (N) y fsforo (F). Se localizan
en todas las clulas. Controlan y coordinan la transmisin de
la informacin gentica y traducen esta informacin para
realizar la sntesis de protenas especficas para cada tipo de
clula.
Existen dos tipos de cidos nucleicos: el cido ribonucleico
(RNA) y el cido desoxirribonucleico (DNA). En ambos
casos los cidos nucleicos estn constituidos por un
conjunto de molculas denominadas nucletidos.
La estructura qumica de un nucletido es la siguiente:
Grupo fosfato + azcar de cinco carbonos (pentosa) + una
base nitrogenada
Acido desoxirribonucleico (DNA). Se sita principalmente
en el ncleo de las clulas. Tambin se le encuentra
integrando a organelos celulares como los cloroplastos
(clulas vegetales) y las mitocondrias (clulas vegetales y
animales). Su estructura qumica es la siguiente:
Grupo fosfato + desoxirribosa + base nitrogenada.
La base nitrogenada puede ser una base pirimdica (citosina
o timina) o una prica adenina o guanina.
El cido desoxirribonucleico contiene la informacin
gentica en su estructura y trasmite los caracteres
hereditarios de una clula a otra.
En 1953, los investigadores J. Watson y F. Crick,
propusieron la teora que la molcula del DNA tiene la
forma de una doble hlice, como una espiral o una escalera
de caracol. Esta configuracin se debe a que las bases
nitrogenadas se unen, una a la otra en forma
complementaria, mediante enlaces de puentes de hidrgeno.
As se unen la base prica adenina (A) con la base
pirimdica timina (T), y la base prica guanina (G) con la
base pirimdica citosina (C).
La molcula de DNA es la nica que se autoduplica debido
a su estructura de doble hlice; una de ellas sirve de plantilla
que determinar la secuencia de las bases de la nueva hebra
o hlice complementaria que se formar. De esta manera la
nueva molcula de DNA resulta idntica a la original.
La molcula de cido desoxirribonucleico es nica y
especfica para cada uno de los organismos animales y
vegetales.
El RNA mensajero transporta la informacin gentica
de sntesis de protenas hacia el citoplasma para
ponerla en contacto con el RNA ribosomal. El RNA de
transferencia conduce los aminocidos necesarios
hacia los ribosomas (RNAr) para el enlace de los
mismos y formar pptidos.
ORGANELOS CITOPLASMTICOS
En el citoplasma se encuentran suspendidos una gran
variedad de componentes figurados, los organelos y las
inclusiones.
Los organelos, denominados tambin organitos, organelas,
u organoides (la mayora de ellos constituidos por
membranas) desempean una variedad de funciones
especficas que intervienen activamente en el metabolismo
celular. Las inclusiones citoplasmticas comprenden a las
sustancias que generalmente almacenan energa, como el
glucgeno y los lpidos y una serie de pigmentos de origen
exgeno o endgeno.
Organelos citoplasmticos
Se clasifican en
membranosos
no membranosos.
Se consideran as a
- Retculo endoplsmico
Figura Biol. Cel. 22. Representacin molecular de la doble hlice del
DNA.
cido ribonucleico (RNA). Tambin es una molcula
compleja pero se diferencia del DNA, est constituido por
una sola hlice o cadena. Forma parte del ncleo y existe en
una buena cantidad en el citoplasma de las clulas.
Su composicin qumica es la siguiente:
Grupo fosfato + ribosa + base nitrogenada
La base nitrogenada puede ser una base prica: la citosina o
el uracilo; o una base pirimdica: la adenina o la guanina.
En las clulas se han identificado tres tipos de RNA,
denominados: ribosomal (RNAr), mensajero (RNAm) y de
transferencia (RNAt). Cada uno de ellos desempea
funciones especficas, todas ellas orientadas a la sntesis de
protenas:
granular o rugoso(RER)
- Retculo endoplsmico
agranular o liso (REL).
- Aparato o complejo de Golgi.
- Mitocondrias.
- Lisosomas.
- Peroxisomas.
- Centriolos, centros celulares
o centrosomas.
- Ribosomas
- Microfilamentos.
- Microtbulos.
- Proteosomas.
RETCULO
ENDOPLSMICO
RUGOSO
O
GRANULAR.
Desde que se iniciaron los estudios de las clulas a travs
del microscopio fotnico y se diferenciaron ciertos
componentes celulares mediante la utilizacin de colorantes,
se pudo observar que muchos tipos celulares posean en el
citoplasma sustancias que se tean de un color intenso con
los colorantes bsicos: hematoxilina, azul de metileno,
fucsina bsica, tionina, azul de toluidina etc. En algunas
clulas esta sustancia le confiere una basofilia difusa al
citoplasma por ejemplo en las clulas hepticas,
condrocitos, eritroblastos y clulas plasmticas o a ciertas
regiones ms o menos precisas del
contenido
citoplasmtico, como se observa en la regin basal de
osteoblastos, de las clulas acinares pancreticas o en forma
de grumos o sustancia Nissl en las neuronas. A este material
basfilo se le denomin ergastoplasma (gr. ergon: trabajo),
o sustancia cromidial, por el color que le confiere a la zona
citoplasmtica o de manera especfica: sustancia Nissl en las
clulas nerviosas.
Las cisternas del retculo endoplsmico rugoso se disponen
de manera paralela, aunque pueden observarse aisladas. Este
tipo de RER existe en aquellas clulas encargadas de
sintetizan protenas cuyo destino es incorporarse a la
membrana plasmtica o que constituirn sustancias de
secrecin (de exportacin). Los ribosomas son los organelos
responsables de la sntesis de protenas.
En otros tipos celulares, especialmente aquellas que
sintetizan protenas estructurales o funcionales que no sern
secretadas, es decir que se quedarn en el citosol; la
basofilia del citoplasma se debe a la presencia de los
polirribosomas o polisomas libres, estructuras integradas
por abundantes ribosomas unidos entre s, mediante una
hebra de RNA mensajero, para formar estructuras que con el
M.E. se observan con una forma levemente espiralada.
Figura Biol. Cel. 24. Expresin morfolgica de la presencia del RER
(retculo endoplsmico rugoso) en clulas observadas con el microscopio
a) sustancia Nissl en neuronas motoras de la mdula espinal, b) basofilia
basal en clulas acinares del pncreas.
Estudios realizados posteriormente demostraron que la
basofilia del citoplasma se deba a la presencia del cido
ribonucleico, pues absorbe la radiacin ultravioleta de 260
nm y la basofilia desaparece cuando las clulas son tratadas
con la enzima ribonucleasa.
Utilizando el microscopio electrnico Keith Porter, en 1945,
observ que en los lugares donde exista basofilia o
sustancia Nissl, se visualizaban una serie de sacos aplanados
o vesculas limitados por membrana; les denomin retculo
endoplsmico porque los sacos aplanados, llamados
cisternas, forman una red anastomosada tridimensional.
Figura Biol. Cel. 25. Dibujo y fotografa electrnica del RER que
muestran la disposicin de las cisternas membranales y los ribosomas
adosados a ella.
La basofilia se debe a la presencia de una gran cantidad de
pequeas partculas de ribonucleoprotenas, que miden 25
nm de dimetro aproximadamente, conocidas como
ribosomas; stos se adhieren a las superficies externas de las
cisternas aplanadas de all el nombre de estos organelos:
retculo endoplsmico rugoso o granular (RER).
Figura Biol. Cel. 26. Imagen ultraestructural de polirribosomas libres.
Las cisternas del retculo endoplsmico rugoso de
disposicin paralela, poseen un espacio interno en forma de
hendidura estrecha que se comunica con el espacio de las
cisternas vecinas mediante anastomosis de las membranas
(fig. Biol. Cel. ). El RER se relaciona directamente con la
membrana externa de la cubierta nuclear y a veces se
relaciona con los componentes membranales del retculo
endoplsmico liso.
A semejanza del plasmalema, las membranas del retculo
endoplsmico estn formadas por tres lminas, dos externas
electrondensas y una media electronlcida. Es posible que
las cisternas se encuentren distendidas cuando la sntesis de
protenas es copiosa y abundante.
Retculo endoplsmico liso (REL) o agranular.
Los componentes membranosos del REL son de aspecto
tubular e integran un sistema anastomosado de tuberias que
se encorvan y en los microambientes citoplasmticos en los
que se localizan. Se ha observado que estos tbulos suelen
mantener comunicacin con las cisternas del RER.
El retculo endoplsmico liso est bastante desarrollado en
las clulas del tejido muscular estriado esqueltico y
cardiaco (retculo sarcoplsmico), de los tbulos renales,
del hgado de las glndulas endocrinas secretoras de
esteroides ( de la corteza suprarrenal, cuerpo amarillo del
ovario y clulas intersticiales o de Leydig del testculo). La
presencia de abundantes tbulos del REL, le confiere
acidofilia al citoplasma que lo contiene.
Figura Biol. Cel. 27. Fotomicrografa del REL
Las funciones que desarrolla el REL son:
Sntesis de lpidos y de hormonas esteroideas. En los
tbulos del REL de los hepatocitos se
sintetizan
colesterol y lipoprotenas. En las clulas adiposas y en las
clulas intestinales de absorcin se sintetizan triglicridos
a partir de los nutrimentos absorbidos. La sntesis de
esteroides se genera en las clulas endocrinas de la
glndula suprarrenal, ovario y testculo endocrinos.
Almacenamiento de glucgeno. Se produce en clulas
que, por su actividad, requieren mantener en reserva
grandes cantidades de energa. En las clulas hepticas y
en las clulas musculares estriadas el glucgeno se
encuentra almacenado como pequeos grumos adheridos
a la superficie membranal externa del REL.
La glucosa se libera a partir de la glucosa-6-fosfato
mediante la enzima glucosa-6-fosfatasa que forma parte
del contenido de protenas de la membrana del REL. La
glucosa fosforilada no puede atravesar la membrana
plasmtica por lo que la glucosa-6-fosfato membranal del
REL elimina el grupo fosfato y genera molculas de
glucosa que en estas condiciones se desplazan hacia la
sangre y de all a otras clulas del organismo.
Desintoxicacin de compuestos orgnicos. Una serie de
sustancias endgenas (catabolitos) y exgenos
(barbitricos y alcohol etlico o pesticidas) son
desintoxicadas por enzimas localizadas en la membrana
del REL. La desintoxicacin se efecta mediante un
conjunto de enzimas que tienen la capacidad de transferir
oxgeno (oxigenasas) por ejemplo el citocromo P450.
Estas enzimas inespecficas tienen la capacidad de oxidar
una gran cantidad de sustancias hidrfobas y convertirlas
en hidrfilas, fciles de excretar.
Secuestro y almacenamiento de iones de Calcio (Ca2+).
En determinadas clulas como las musculares estriadas
(esquelticas y cardiacas) el REL, se dispone en la forma
de retculo sarcoplsmico en cuyo interior se capturan y
se regula la liberacin de iones de Ca2+. En las fibras
musculares, los iones de calcio desempean un papel
sumamente importante en la contraccin muscular.
Sntesis de los lpidos de la membrana celular. En las
clulas eucariotas la sntesis de los fosfolpidos de la
membrana celular se produce en el REL, especficamente
en la hoja citoplasmtica de la estructura trilaminar de los
tbulos. Posteriormente se desplazan a la capa interna ya
sea utilizando procesos enzimticos o a travs de
movimientos de balanceo o de flip flop. Una vez
instalados los fosfolpidos en la membrana interna de los
tbulos son conducidos al plasmalema siguiendo el
mismo recorrido que efectan las protenas membranales
e incorporarse a la bicapa de fosfolpidos; esto significa
que del REL, los fosfolpidos deben continuar con las
cisternas del RER, las vesculas de transferencia, las
diversas cisternas del aparato de Golgi y, a travs de
pequeas vesculas secretorias adherirse a la membrana
celular. Para tal fin suelen emplear las denominadas
protenas transportadoras de fosfolpidos que transfieren
estas molculas de una membrana celular a otra.
Aparato o complejo de Golgi.
El aparato de Golgi fue observado por primera vez en 1898,
por Camilo Golgi, bilogo italiano mediante el uso de
tcnicas de impregnaciones metlicas como las sales plata,
de osmio y de otros metales pesados. Golgi observ en
neuronas de animales, despus de impregnar el tejido, una
especie de retculado pardo oscuro que ocupaba la casi
totalidad del citoplasma alrededor del ncleo. Esta imagen
fue designada por Golgi con el nombre de aparato reticular
interno. Aos ms tarde se le llam Aparato de Golgi, en
honor a su descubridor
Gracias a sus hallazgos citolgicos y a sus aportes a la
citoqumica y la histoqumica, Camilo Golgi, junto con el
mdico espaol Santiago Ramn y Cajal, fueron
galardoneados en 1906, con el premio Nobel.
Figura Biol. Cel. 28. Imgenes fotogrficas del aparato de Golgi, en
clulas nerviosas ganglionares (posicin perinuclear) y en clulas
epididimarias (posicin supranuclear) TGN
Durante muchos aos la existencia de este organelo fue
motivo de controversias cientficas porque, debido a la
dificultad de demostrarlo mediante las tcnicas de
impregnaciones metlicas, cuyos resultados no siempre eran
fructferos, se le consider como un artefacto de tcnica. El
aparato de Golgi no puede demostrarse cuando las clulas se
tien con colorantes derivados de la anilina, por ejemplo
hematoxilina eosina (HE), u otras tcnicas que no son de
impregnacin metlica. En estos casos, se le observa como
una zona no coloreada, imagen negativa del aparato de
Golgi, generalmente relacionada con el ncleo; rodeada
por una basofilia difusa, especialmente en aquellas clulas
que desarrollan funciones de sntesis y secrecin de
protenas como las clulas plasmticas o los osteoblastos.
convexa, en la parte inferior, denominada superficie
inmadura, de formacin, cara o superficie cis es la ms
cercana a la cubierta nuclear y al RER, la superficie
superior muestra una curvatura cncava, se le conoce con el
nombre de superficie o cara madura, o superficie trans. Esta
superficie se localiza en el polo opuesto y est ms cercana
a la membrana celular. En medio de estos grupos de
cisternas, se sitan otras, conocidas como porcin
intermedia o medial.
En la dcada de 1950, con el empleo del microscopio
electrnico se confirm la existencia del aparato de Golgi
considerndolo como un organelo celular. Los estudios de
su ultraestructura sirvieron para incrementar sus detalles
morfolgicos.
Estudios realizados para identificar enzimas en las cisternas
del aparato de Golgi, demuestran que las cisternas que lo
constituyen no tienen una estructura molecular uniforme;
por ejemplo, el tetraxido de osmio, impregna de manera
selectiva las dos o tres cisternas de la superficie cis. Las
cisternas de la cara intermedia o medial dan reaccin
positiva a la enzima manosidasa II, encargada de eliminar
los residuos de manosa en tanto que, en la superficie trans se
demuestra la presencia de la enzima difosfatasa de
nuclesido, responsable de desdoblar dinucletidos.
Con las tcnicas de impregnacin metlica, especialmente
aquellas que utilizan sales de plata, el aparato de Golgi se
muestra de dos formas, en distintos tipos celulares, en todos
los casos siempre relacionado con el ncleo.
En las neuronas y en los hepatocitos, los componentes
reticulares aparecen alrededor del ncleo (posicin
perinuclear), en cambio en otras clulas que exhiben cierta
polaridad funcional secretora, por ejemplo en las clulas
acinares del pncreas, clulas caliciformes, clulas
plasmticas o las del epiddimo, el aparato de Golgi se
observa como un pequeo retculo en forma de ovillo
situado encima del ncleo (posicin supranuclear), en el
tercio apical.
Figura Biol. Cel. 30. Esquema representativo de las cisternas y vesculas
que integran el RECIG y RGT del aparato de Golgi.
Figura Biol. Cel. 29. Representacin esquemtica y fotomicrografa del
aparato de Golgi, observado mediante el M E. T.
En la actualidad el aparato o complejo de Golgi se describe
como un conjunto de cisternas ovales aplanadas (se semejan
a un plato) colocadas unas encimas de las otras formando
pilas (fig. Biol. Cel. ), a este grupo de cisternas tambin se
les denomina dictiosomas; rodeadas en su circunferencia, a
los lados, abajo y arriba de ellas por vesculas de diverso
dimetro
31/07/03
El nmero de cisternas de Golgi en cada dictiosoma es de 5
a 10, la luz de las cisternas mide de 500 a 1000 nm de
grosor. Los bordes de cada cisterna estn ligeramente
curvados y muestran dilataciones y perforaciones en forma
de fenestras. Esta disposicin le confiere a cada aparato de
Golgi la forma de platillos dispuestos con una curvatura
Al conjunto de cisternas se les aaden dos compartimentos
adicionales, uno relacionado con la cara cis y otro con la
cara trans. Entre la cara cis y el RER se ubica un conjunto
de vesculas denominado retculo endoplsmico rugosocompartimento intermedio de Golgi (RECIG) y, la red de
Golgi trans (RGT), localizado sobre la cara trans. El
RECIG es un conjunto de vesculas y tbulos constituidos
por la fusin de las vesculas de transferencia o de
transporte provenientes de la porcin final del RER, que
contienen las protenas recientes sintetizadas en el RER. El
trnsito de las protenas sintetizadas sigue un recorrido que
se inicia con las vesculas de transferencia o de transporte
desprendidas de las cisternas finales del RER, ahora
carentes de ribosomas (retculo endoplsmico transicional),
stas se incorporan al RECIG para fusionarse
posteriormente con la superficie o cara cis del aparato de
Golgi con la finalidad que las protenas transportadas
puedan modificar su estructura molecular. Las protenas son
conducidas mediante vesculas o anastomosis membranales
que se generan en las ltimas cisternas de la cara cis a las
cisternas de la cara medial y posteriormente, siguiendo el
mismo patrn de transporte lleguen a la superficie trans.
Conforme las protenas discurren a travs de las cisternas
del aparato de Golgi, van siendo modificadas, por ejemplo
las protenas que constituirn glucoprotenas se glucosilan
de manera notable, en tanto que otras pierden o adquieren
ciertas molculas de carbohidratos.
La fosforilacin de la manosa se lleva a cabo en las cisternas
de la cara cis, en tanto que la prdida de la manosa de otras
protenas se realiza en las cisternas de la cara cis y de la
porcin medial. En las cisternas mediales se aaden a las
protenas el azcar N-acetilglucosamina, en cambio la
adicin del cido silico (cido N-acetilneuramnico) y de
galactosa, as como la fosforilacin y sulfatacin de
aminocidos se produce en las cisternas de la superficie
trans del aparato de Golgi.
VESCULAS RELACIONADAS CON EL RER Y EL
APARATO DE GOLGI.
Las vesculas que conducen protenas sintetizadas en el
RER hacia el aparato de Golgi y las vesculas que emergen
de la superficie trans de este organelo para incorporarse a la
membrana celular o a otros organelos requieren del marcaje
de cubiertas protenicas del lado del citoplasma para
emerger del RER o que estas cubiertas les indiquen el
camino para trasladarse de un organelo a otro.
Figura Biol. Cel. 32. Caractersticas estructurales de las vesculas
relacionadas con el RER y el aparato de Golgi.
Las cubiertas protenicas son de tres tipos: coatmero I
(COP I), coatmero II (COP II), a las cuales se les denomina
protenas residentes, y clatrina. El proceso de
desprendimiento de las vesculas se inicia en las cisternas
del RE libres de ribosomas con la cubierta de coatmero II
(COP II), las vesculas as cubiertas se insertan en el
RECIG; en este lugar el contenido de las vesculas se libera
para continuar su trayecto hacia la cara cis, en tanto que las
protenas residentes integrantes del COP II, se reciclan en
forma de pequeas vesculas se reciclan y vuelven al RER.
A este tipo de transporte se le conoce como transporte
retrgrado. Las vesculas conteniendo las protenas
sintetizadas emergen del RECIG para incorporarse a la
superficie cis y de estas cisternas a las mediales, luego las
cisternas de la cara trans y de all a la red de Golgi trans
(RGT). Estas vesculas requieren de la presencia de la
cubierta protenica coatmero I (COP I). Este tipo de
transporte de avance desde el RER, hasta la cara CIS del
aparato de Golgi se denomina transporte antergrado.
La mayora de las vesculas que provienen de la red de
Golgi trans (RGT) necesitan de la protena clatrina para su
formacin y transporte.
Una vez liberadas las vesculas conteniendo protenas
modificadas, es necesario que se trasladen a diversos
lugares de la clula; en ciertos casos la membrana de la
vescula posee marcadores o receptores de superficie, los
cuales interaccionan con microtbulos y complejos de
protenas motoras que se encargan del movimiento y
traslado de las vesculas.
02/07/03. 16:15 horas
Las molculas motoras de los microtbulos son dos: la
dinena y la cinesina. La dinena hace avanzar las vesculas
hacia el polo terminal de los microtbulos esto significa que
interviene en el transporte antergrado, en cambio la
cinesina conduce las vesculas de reciclaje hacia el polo
inicial de los microtbulos, ejerciendo su capacidad motora
en el transporte retrgrado de vuelta al RECIG o al RER.
SELECCIN DEL DESTINO DE LAS PROTENAS
EN LA RED DE GOLGI DE LA CARA TRANS
(RGT).
Las sustancias protenicas emergen del RGT dentro de
vesculas membranales constituidas por protenas
especficas pertenecientes a la familia de protenas vSNARE codificadas por genes especficos para integrar
vesculas de transporte. Otras protenas insertas en las
membranas aceptoras especficas
que reciben a las
vesculas de transporte, son codificadas por genes que
expresan las protenas de la familia t-SNARE. La
interaccin especfica entre las protenas v-SNARE y las tSNARE coordinan el encuentro y la fusin de las
membranas de las vesculas transportadores y las
membranas de los componentes celulares aceptores.
El destino del contenido de las vesculas originadas en el
RER y que discurren a travs del aparato de Golgi es.
a) Insertarse en el plasmalema como protenas y lpidos de
la membrana.
b) Mediante la fusin de la membrana vesicular con el
plasmalema el contenido se vierte inmediatamente hacia
el espacio extracelular (secrecin constitutiva).
c) Acumularse en el citoplasma cercano al plasmalema, en
la forma de vesculas voluminosas conocidas vesculas
de condensacin o tambin como grnulos de
secrecin y, mediante una seal (estmulo) especfica,
fusionarse con la membrana celular para la liberacin
final (secrecin) del contenido hacia fuera de la clula
(secrecin regulada).
d) Marcarse mediante fosforilacin, con manosa-6-fosfato
y de esa manera identificarse en la forma de protenas
enzimticas lisosomales con la finalidad que sean
seleccionadas para incorporarse como vesculas que
emergen del RGT y originar los lisosomas.
Secrecin constitutiva. Es propia de la mayora de las
clulas; se caracteriza porque representa un proceso
continuo de sntesis en el RER, modificaciones moleculares
en el Aparato de Golgi, formacin de pequeas vesculas,
las cuales sin necesidad de seales o estmulos especficos,
fusionan sus membranas con el plasmalema y vierten su
contenido al exterior de la clula. La secrecin constitutiva
se encarga de secretar (exocitosis) el producto contenido en
las vesculas de sustancias como factores de crecimiento,
molculas que integran la sustancia intercelular: membranas
basales, matriz amorfa, glucosaminoglucanos, fibras
colgenas y elsticas y enzimas. Tambin es el proceso que
propicia el suministro de componentes lipdicos, protenicos
y de carbohidratos a la membrana celular. Durante este
proceso las vesculas se recubren de coatmeros I (COP I),
es decir cubiertas protenicas que no le confieren una
seleccin especfica al contenido de las vesculas.
Secrecin regulada. Se produce en clulas especializadas
en la secrecin de sustancias especficas. Por ejemplo, en las
clulas de los acinos pancreticos las vesculas se acumulan
en la regin apical de la clula (entre el ncleo y el
plasmalema) en la forma de grnulos de secrecin que
muestran dimetros fcilmente visibles con el microscopio
fotnico; la fusin de las membranas vesiculares con la
membrana celular para generar exocitosis, slo se produce
como respuesta seales o estmulos especficos (hormonales
o neurotransmisores) o en vesculas que contienen protenas
enzimticas especficas destinadas a convertirse en
lisosomas.
Durante la emergencia de las vesculas de la cara trans o del
RGT del aparato de Golgi, las vesculas se cubren de la
protena clatrina. Se considera que la clatrina contribuye al
proceso de gemacin y separacin de las vesculas del RGT.
Los componentes de la clatrina, denominados trisquiliones
de clatrina, se separan inmediatamente despus que los
grnulos se desprenden del aparato de Golgi.
Los trisquiliones de catrina son molculas que se encuentran
suspendidos en el citoplasma. Ante la necesidad de
recubrimiento que requieren ciertas vesculas, para liberarse
del RGT, los trisquiliones se ensamblan y constituyen una
cubierta en forma de canasta o cesto que rodea ntegramente
a las vesculas. En ellas ejercen presin (una especie de
pellizco) sobre la unin que todava las sujeta a la
membrana del RGT que las origin, hasta que la vescula se
separa totalmente de su unin membranal.
NCLEO.
Es el organelo de mayor tamao de la clula. Alberga la casi
totalidad
del
material
gentico,
el
cido
desoxirribonucleico (DNA) de la clula; componente bsico
e indispensable para la realizacin de la mayora de las
funciones celulares y la transmisin de los caracteres
hereditarios. Mediante otro componente nuclear, el cido
ribonucleico (RNA), cumple los mecanismos necesarios
para la sntesis de protenas. La gran mayora de clulas
eucariotas que constituyen un organismo pluricelular poseen
ncleos. Los eritrocitos de los mamferos y algunas clulas
terminales de una estirpe tisular, por ejemplo las clulas del
estrato crneo de la epidermis, carecen de ellos, porque son
clulas que durante su ciclo vital han perdido la capacidad
de sintetizar protenas y de multiplicarse.
Morfologa general del ncleo.
La forma del ncleo vara dependiendo de la forma que
presenta la clula. Generalmente es de forma esfrica en
clulas redondeadas, polidricas o cbicas. Adoptan una
forma alargada u oblonga en las clulas cilndricas,
fusiformes, siguiendo el eje de mayor longitud de las
clulas. En clulas sanguneas leucocticas suelen mostrar
formas lobuladas (granulocitos) o con ciertas escotaduras
(monocitos) ms o menos profundas.
SHAPE
\*
MERGEFORMAT
a) La envoltura nuclear o nucleolema, representada por
una fina lnea coloreada que limita el contenido del
ncleo del citoplasma.
b) La cromatina, se distingue como un conjunto de
grnulos o grumos irregulares de color basfilo intenso,
dispuestos sobre la superficie interna de la cubierta
nuclear o dispersos en el interior del ncleo. La
cromatina se dispone de dos formas, los grumos teidos
intensamente corresponden al enpaquetamiento o
condensacin del DNA por la presencia de protenas,
denominadas histonas. A este tipo de disposicin se le
denomina cromatina condensada o heterocromatina
(DNA que no expresa rdenes para la sntesis de
protenas) y la disposicin de la cromatina extendida o
no empaquetada, no coloreada, es la cromatina en la cual
el DNA se encuentra transcribiendo rdenes para la
sntesis protenica. Se le conoce como eucromatina o
cromatina laxa, no visible con el microscopio fotnico.
c) Uno o ms nuclolos, estructuras esfricas u ovaladas
constituidos principalmente por ribonucleoprotenas
donde predomina el cido ribonucleico y
d) El jugo nuclear o carioplasma, constituido
principalmente por agua, electrolitos y iones.
Figura Biol. Cel. 33. Formas y tamaos de ncleos pertenecientes a varios
tipos de clulas animales, observados a travs del microscopio fotnico.
El tamao del ncleo es variable, depende del tipo celular,
casi siempre es ms grande en las clulas ms voluminosas
que en las clulas de menor tamao; en las clulas de
mamferos mide entre 5 y 10 micrmetros.
La cantidad de ncleos en el interior de las clulas tambin
puede variar; generalmente existe un solo ncleo en cada
clula, pero existen clulas que pueden poseer ms de un
ncleo. Algunas clulas hepticas y de la corteza
suprarrenal, como ciertas neuronas de ganglios raqudeos
suelen albergar dos ncleos, son binucleadas. Otras clulas
como los osteoclastos, las fibras musculares estriadas
esquelticas tienen varios ncleos, son clulas
multinucleadas.
Componentes del ncleo. Cuando se observa una clula
eucariota coloreada con hematoxilina eosina, el ncleo se
tie con el colorante bsico, la hematoxilina, el color vara
de azul, morado, pardo o negro. Con el microscopio
fotnico la imagen del ncleo nos muestra los componentes
siguientes:
Figura Biol. Cel. 34. Componentes del ncleo observados mediante los
microscopios fotnico y electrnico.
04/07/03 20:30 horas
Envoltura nuclear o cariolema..
Es un componente del ncleo que marca la diferencia entre
las clulas eucariotas de las procariotas, el material gentico
de las eucariotas se encuentra rodeado de una fina
membrana, distinguible con el microscopio fotnico, que lo
separa del citoplasma. Fue observada por primera vez por
Hertwig en 1893.
La envoltura nuclear, examinada mediante microscopa
electrnica, est constituida por dos membranas celulares,
una interna y otra externa, existiendo entre ambas por
espacio denominado cisterna perinuclear. Cada cierto
trecho la cubierta muestra perforaciones llamadas poros
nucleares en donde las dos membranas se fusionan entre s
para permitir la comunicacin entre el citoplasma y el
nucleoplasma. A travs de los poros nucleares, la envoltura
nuclear coordina el transporte de macromolculas entre el
citoplasma y el ncleo. Debajo de la membrana interna del
ncleo se dispone la lmina densa,
formada por un
conjunto de protenas fibrilares encargada de proporcionarle
soporte a la cubierta nuclear y servir como zona de anclaje o
de fijacin para la organizacin de la heterocromatina
perifrica.
La membrana nuclear externa mide aproximadamente 6nm
de grosor, su superficie externa se orienta hacia el
citoplasma, en ella se adhieren ribosomas, lo que le
proporciona una imagen similar al RER. Los ribosomas se
encargan de la sntesis de protenas transmembranales de las
dos membranas de la cubierta nuclear. Cada cierto trecho se
observa la continuidad de la cubierta nuclear con el retculo
endoplsmico rugoso perinuclear. En su superficie externa
se visualiza una malla laxa y fina constituida por filamentos
intermedios de vimentina.
La membrana nuclear interna, tambin tiene un grosor de
unos 6 nm, est en contacto estrecho con el carioplasma y la
lmina densa nuclear, integrada por una malla densa de
filamentos intermedios integrados por polmeros de
polipptidos denominados lminas, de 80 a 100nm de
espesor. Las lminas son de tres tipos A, B y C. Las lminas
tipo A se sitan en la superficie citoplasmtica de la lmina,
las del tipo B predominan en la superficie de la lmina que
est en contacto con la membrana interna y establecen
relacin estrecha con protenas integrales de aquella. Sirven
como zona de sujecin o soporte a la cromatina condensada
en las clulas en interfase; durante la mitosis se postula que
sirve para orientar a los cromosomas especialmente en las
dos primeras fases de la divisin celular. En la regin de los
poros nucleares se percibe la ausencia de la lmina densa y
de la heterocromatina.
Figura Biol. Cel. 35. Imagen de la ultraestructura de los componentes de la
cubierta nuclear.
A semejanza de los filamentos intermedios del citoplasma,
la integridad de la lmina densa nuclear esta mediada por la
fosforilacin o desfosforilacin de las protenas que la
integran. Se considera que la desintegracin o
desensamblado de la lmina nuclear antes de mitosis ( etapa
final de la G2) es provocada por fosforilacin de las lminas
mediante cinasas.
Poros nucleares. Son aberturas constituidas por la fusin de
las membranas nucleares externa e interna localizadas cada
cierto trecho de la cubierta nuclear. Los poros nucleares
facilitan la comunicacin del citoplasma con el contenido
nuclear.
La distribucin de los poros no es regular en la cubierta
nuclear de una clula y existen variaciones dependiendo del
tipo de clula. Se calcula que el nmero de poros dispuestos
en la cubierta nuclear vara desde 5 a 60 poros por
micrmetro cuadrado. Los poros funcionan como puertas
que dejan pasar en uno u otro sentido una apreciable
cantidad de macromolculas, el RNA y una variedad de
protenas utilizan los poros para trasladarse entre ambos
microambientes.
Figura Biol. Cel. 36. Fotomicrografa electrnica de la cubierta nuclear
mostrando los poros nucleares, obtenida mediante el procedimiento de
criofractura.
Para que el material gentico DNA y RNA se duplique es
necesario que una serie de componentes que los integran,
discurran del citoplasma hacia el ncleo y, del interior del
ncleo hacia el citoplasma deben transportarse, atravesando
la cubierta nuclear, las subunidades ribosomales, el RNA
mensajero y el RNA de transferencia sintetizados en el
ncleo.
Podra pensarse que los poros nucleares, al dejar pasar con
relativa facilidad estas macromolculas actuaran
simplemente como canales abiertos, pero esto no es as, los
poros nucleares contienen una conformacin sumamente
compleja, en forma de canastilla tanto que algunos autores
las designan como complejo de poro nuclear (CPN) que se
inserta como un tapn en el espacio del poro y sobresale
ligeramente hacia el citoplasma y hacia el nucleoplasma.
El complejo de poro nuclear esta formado por una estructura
octogonal cilndrica de 120 nm de dimetro. En el extremo
citoplasmtico, el poro consta de un anillo externo, formado
por ocho ovillos protenicos que se disponen en forma
circular, de estos se proyectan al citoplasma filamentos de
protenas, y por el lado del nucleosoma posee un anillo
interno, tambin constituido por ocho ovillos de disposicin
circular. Ambos anillos estn unidos entre s por columnas
protenicas que se disponen en forma vertical. De las
columnas se proyectan puentes radiales (constituidos por
glucoprotenas) que se unen en el centro del complejo a un
grnulo central conocido como transportador central, ste
tiene la forma de un reloj de arena, y posee en el centro un
canal axial que mide aproximadamente unos 8 a 10 nm de
dimetro (fig. Biol.Cel.). Del anillo interno pende hacia el
nucleoplasma, una estructura filamentosa flexible, en forma
de canasta, la canastilla nuclear.
produce un cambio de conformacin del transportador
central el cual se abre formando un canal acuoso. El traslado
de ribonucleoprotenas del ncleo al citoplasma debe
implicar acciones semejantes. El paso de molculas del
citoplasma al nucleoplasma y viceversa, se produce por un
mismo poro. Las protenas que intervienen en incorporar
macromolculas al interior del ncleo se denominan
importinas; aquellas que sacan ribonucleoprotenas hacia el
citoplasma se llaman exportinas.
Cromatina. Es el contenido nuclear que se tie con
colorantes bsicos. Esta integrada por cadenas de DNA y
protenas asociadas; histonas y no histonas. Las histonas son
un conjunto de protenas agrupadas en cinco grupos que se
diferencian entre ellas por su contenido de los aminocidos
arginina y lisina. Las histonas se designan como H1, H2A,
H2B, H3 y H4. Las protenas no histonas son protenas
enzimticas reguladoras de estructura muy diversa.
Las histonas H1, H2A y H2B son ricas en lisina. La
presencia del aminocido lisina permite a estas histonas
tener una mayor afinidad por el DNA.
Figura Biol. Cel. 37. Fotomicrografa electrnica y esquema del poro
nuclear y las estructuras que lo constituyen.
El transporte de partculas a travs de la cubierta nuclear se
efecta mediante varios procedimientos: Molculas
pequeas y iones atraviesan con rapidez ambas membranas,
molculas, de mayor tamao y peso molecular discurren a
travs de las membranas en forma de pequeas vesculas
que atraviesan las dos membranas y liberan su contenido al
otro lado y, por ltimo el paso de sustancias se realiza
atravesando el poro nuclear. El transporte se realiza
mediante dos mecanismos. Las molculas menores a 10 nm
atraviesan los poros por difusin pasiva, entre los rayos de
la canastilla. Molculas de mayor tamao utilizan
mecanismos que requieren de gasto de algn tipo de
energa.
Las protenas que no integran componentes nucleares no
atraviesan el poro, en cambio protenas que son residentes
del ncleo, como la nucleoplasmina (abundante en ovocitos
de anfibios) pasan fcilmente a travs del poro. Los estudios
realizados, en 1982; por Robert Laskey y colaboradores, en
Inglaterra, demostraron que la nucleoplasmina posee un
tramo de aminocidos de carga positiva que acta como
seal de localizacin nuclear (SLN); la seal le permite
unirse a receptores especficos del anillo externo del poro
nuclear. Los receptores especficos residen en las protenas
filamentosas relacionadas con el citoplasma. De esa manera
las protenas nucleares (sintetizadas previamente en el
citoplasma) pueden incorporarse al interior del ncleo. El
desplazamiento de la protena a travs del poro requiere del
empleo de energa que se obtiene por hidrlisis de ATP, esto
El nombre de cromatina se debe a la facilidad con que se
tien intensamente. Es la misma reaccin de coloracin que
poseen los cromosomas (cadenas de DNA enrolladas de
manera sumamente compacta) en las fases de la mitosis y de
la meiosis. La cromatina se dispone en el interior del ncleo
de dos formas, condensada como grumos irregulares
intensamente coloreados que en conjunto conforman la
heterocromatina y la dispuesta de manera laxa, que ocupa
los espacios aparentemente vaco del nucleoplasma, se le
denomina eucromatina.
No todo el material basfilo del ncleo est constituido por
desoxirribonucleoprotenas, por ejemplo el cido
ribonucleico, que forma los principales componentes del
nuclolo y las unidades que conforman los ribosomas.
Existe una reaccin histoqumica desarrollada por Feulgen
que identifica especficamente al DNA, colorendolo de
rojo magenta. Los nuclolos ni el contenido ribosomal se
tien.
a)
b)
Figura Biol. Cel. 38. Fotomicrografas de clulas con ncleos de a)
cromatina condensada de linfocitos (ncleos de cara cerrada) y de b)
cromatina laxa de neuronas (ncleos de cara abierta).
Est demostrado que los grumos de cromatina condensada
son las porciones enrolladas de los cromosomas o DNA
inactivo, que no expresa rdenes de sntesis de protenas y
la cromatina laxa son las regiones extendidas de los
cromosomas o DNA activo que exhiben sus nucletidos
para que ellos expresen rdenes de formar RNA mensajero,
con la finalidad de propiciar la sntesis de protenas.
La heterocromatina se dispone dentro del ncleo de tres
maneras:
Cromatina dispersa en el citoplasma como
pequeos grupos enlazados entre s por finos
filamentos irregulares de cromatina laxa.
Cromatina perifrica, localizada como grumos
gruesos y en contacto estrecho con la lmina
nuclear densa y,
Cromatina asociada al nuclolo que rodea a este
componente nuclear y enva ramificaciones
delgadas al interior del mismo..
El patrn de distribucin de la heterocromatina y de la
eucromatina en el interior del ncleo, nos permite deducir su
capacidad de sntesis de protenas de una clula, cualquiera
que sea el tipo de clula que observemos. Cuando existe un
predominio de cromatina densa o compacta en la estructura
del ncleo (ncleo de cara cerrada) como el de los linfocitos
circulantes (fig. Biol. Cel. 38) o clulas de los estratos ms
superficiales de la epidermis o el ncleo de los
espermatozoides, podemos afirmar que esas clulas son
inactivas en la sntesis de protenas, en cambio, los ncleos
que muestran un predominio de cromatina laxa o extendida
(ncleos de cara abierta) como el de los hepatocitos, las
neuronas (fig. 32), de las glndulas exocrinas o endocrinas o
de las clulas plasmticas (linfocitos B activos), debemos
deducir que pertenecen a clulas con gran actividad de
sntesis. Se define a la cromatina como
06/08/03 18:50 horas
Cromosomas. Los cromosomas alcanzan su mxima
compactacin y se observan fcilmente en la metafase
(cadena de DNA duplicada) de la divisin celular. El
nmero de cromosomas que poseen las clulas de una
especie animal o vegetal es variable entre las diferentes
especies pero cada una de ellas posee una determinada
cantidad. Las clulas somticas aquellas que forman tejidos
y rganos, con excepcin de las clulas sexuales o
germinales: ovocitos y espermatozoides constituyen el
genoma, es decir la caracterizacin gentica total de un
organismo, por ejemplo un parsito del aparato
gastrointestinal del caballo, la lombriz Ascaris
megalocephala poseen dos cromosomas, las clulas de la
mosca de la fruta, Drosophila melanogaster tienen 8
cromosomas, las clulas del chimpanc y de la papa, poseen
48 cromosomas, la vaca tiene 60 cromosomas; las clulas de
la especie humana (Homo sapiens sapiens) poseen 46
cromosomas.
Las clulas sexuales o germinales tienen la mitad de los
cromosomas de cada especie.
El genoma contiene toda la informacin gentica o la
herencia de un individuo.
La unidad de herencia de un ser vivo se denomina GEN;
consiste en una secuencia de bases del DNA que contiene la
informacin necesaria para la sntesis de un cido nucleico
o de una protena.
Un
complejo molecular integrado por cido
desoxirribonucleico y protenas del tipo histonas y no
histonas que representan el material gentico compacto o
laxo constitutivo de los cromosomas en el ncleo en
interfase.
Organizacin de la cromatina. En el ncleo en interfase la
cromatina se dispone como eucromatina y heterocromatina.
En el ncleo de la clula que se encuentra en el proceso de
mitosis o divisin celular, la cromatina se organiza en unos
cuerpecillos intensamente teidos que se denominan
cromosomas. stos son visibles mediante el microscopio
fotnico. Los cromosomas son el producto de la
compactacin extrema de las cadenas de DNA + protenas
(desoxirribonucleoprotenas). En 1888, Waldeyer emple el
trmino de cromosoma (cuerpo coloreado) para designar a
estas estructuras, por la afinidad que tienen para teirse
intensamente con colorantes bsicos.
Figura Biol. Cel. 39. Cariotipo de clulas somticas humanas. Se observan
los pares de cromosomas arreglados por la posicin del centrmero y el
tamao de los brazos.
El arreglo secuencial de bases que integran el gen representa
la secuencia de aminocidos de la protena. El cdigo
gentico est constituido de tal manera que un triplete de
bases consecutivas, denominado codn, conforma un
aminocido particular.
En la actualidad el proyecto denominado del Genoma
Humano ha logrado determinar, secuenciar e identificar,
como si fuera un mapa, 100, 000 genes y alrededor de 3000
millones de nucletidos integrantes del genoma de la
especie humana.
Los cromosomas se agrupan por pares, en la especie
humana existen 23 pares de cromosomas. Cada par de
cromosomas se constituye por el aporte, durante la
fecundacin, de cromosomas similares en su contenido
gentico y en su estructura morfolgica, proporcionados por
las clulas germinales de los padres: 23 cromosomas
aportados por el ovocito y 23 cromosomas por el
espermatozoide. El genoma y las formas de los
cromosomas se heredan de una generacin de clulas a otra
y lo mismo ocurre en todos los organismos.
Los cromosomas (cromatidas) en las clulas en interfase
estn formados por una sola molcula de DNA; cuanto ms
grande sea la molcula de DNA, mayor tamao exhibir el
cromosoma.
En los inicios de la dcada del 70, se demostr que cuando
la cromatina se trata con nucleasas inespecficas se
descompone en fragmentos de aproximadamente 200 pares
bases; en 1974, Roger Kornberg postul que el DNA y las
histonas se organizan en unidades repetitivas llamadas
nucleosomas. stos estn constituidos por a) la partcula
central del nucleosoma, que semeja la cuenta de un collar,
formado por una secuencia de 146 pares de bases que se
enrollan en casi dos vueltas alrededor de b) un complejo de
ocho molculas de histonas; el complejo est constituido
por cuatro pares de las histonas H2A, H2B, H3 y H4; en
conjunto las partculas centrales miden 10 nanmetros de
grosor. Las partculas centrales estn enlazadas entre s a
travs de un segmento de DNA de unin de una longitud de
ms o menos 60 pares de bases con de grosor de 2
nanmetros. En el punto de entrada y salida de la hebra de
DNA se dispone la histona H1. El conjunto de partculas
centrales y el segmento de unin muestran en las
fotomicrografas electrnicas el aspecto de las cuentas de
un rosario.
La disposicin de la cadena de pares de bases y las histonas
en nucleosomas de 10-11 nm de dimetro, representan la
organizacin ms simple del enpaquetamiento de la
cromatina.
Figura Biol. Cel. 39. Esquema que representa la organizacin de un
cromosoma a partir de la doble hlice de DNA y las nucleoprotenas que la
compacta.
Cuando se observan los grumos de cromatina
(heterocromatina) o los filamentos no condensados de la
eucromatina, al microscopio electrnico, se distinguen
porciones en forma de filamentos electrondensos de un
grosor aproximado de 30 nanmetros. Se ha propuesto que
este tipo de disposicin de la cadena de DNA se debe al
enrollamiento que experimentan los nucleosomas sobre s
mismos en una estructura denominada solenoide. Los
solenoides resultan del enpaquetamiento de los nucleosomas
en nmero de seis, gracias a las interacciones de las
molculas de histonas H1, ricas en lisina, que se establecen
con nucleosomas vecinos.
Figura Biol. Cel. 40. Fotomicrografa electrnica de una cadena de
solenoides unidos por la doble hlice de DNA.
Conforme la clula termina la etapa de interfase y se prepara
para la divisin celular, las fibras cromatnicas constituidas
por solenoides, vuelven a enrollarse para formar asas de
300 nm de grosor, que se unen mediante la interaccin de
DNA con protenas. La condensacin se hace ms evidente
cuando en la profase las asas se condensan de manera
espiralada para formar haces helicoidales de 700 nm de
dimetro y compactarse an ms para constituir los brazos
de los cromosomas en metafase (cromatidas duplicadas). Se
calcula que un cromosoma en la metafase mide
aproximadamente 1.4 micrmetros de grosor.
Las diferentes formas de enrollamiento que se han
mencionado permiten reducir en casi 10,000 veces la
longitud total de la cadena de DNA lo que permite
almacenar una gran cantidad de informacin gentica en
compartimentos tan pequeos como son los ncleos,
algunos de los cuales miden aproximadamente unos 5 a 6
micrmetros de dimetro.
se esparza lo suficiente como para formar una delgada
capa de cromosomas distribuidos al azar.
Tipos de cromosomas.
En todo organismo pluricelular existen dos tipos de
cromosomas, los autosomas y los cromosomas sexuales. En
la especie humana existen 44 cromosomas autosomas
agrupados en 22 pares, son los cromosomas que contienen
en su mayor parte el cdigo gentico que gobierna y
coordina la reproduccin, proliferacin y diferenciacin de
casi todas las clulas de los tejidos y rganos; y dos
cromosomas sexuales, reunidos en un par. En la mujer son
los cromosomas XX y en el varn son los cromosomas XY.
Los cromosomas sexuales son los responsables de
establecer el sexo gentico (femenino o masculino) de cada
individuo despus de producida la fecundacin.
Coloracin de los cromosomas. Se fija la preparacin y
los cromosomas se tien empleando diferentes
colorantes: orceina, el colorante de Giemsa para teir
bandas especficas de cromatina (bandas G) que
identifican de manera precisa los pares de cromosomas
homlogos, o se emplean colorantes especiales,
fluorocromos como la quinacrina que mediante el uso
del
microscopio
de fluorescencia
(radiacin
ultravioleta) facilita, de manera especfica, la
identificacin, los diversos tipos de cromosomas
(identificados por las bandas Q).
Caractersticas morfolgicas de los cromosomas
humanos.
Para poder establecer las caractersticas morfolgicas de los
cromosomas, es necesario desarrollar y aplicar ciertos
procedimientos que permiten individualizarlos y conferirles
determinado tipo de marcaje mediante el empleo de tcnicas
de tincin. Los cromosomas alcanzan su mejor caracterstica
morfolgica y de coloracin en la etapa de metafase de la
divisin celular.
La tcnica que se utiliza con ms frecuencia consiste en los
siguientes pasos.
Fotografiado del cariograma. Se denomina cariograma
a la imagen fotogrfica que se obtiene de los
cromosomas coloreados.
Conformacin del cariotipo. Se ampla la fotografa; se
recortan las siluetas de los cromosomas y se les ordena
por pares de acuerdo al tamao, forma y longitud de los
brazos.
Obtencin de clulas. Para tal fin se utilizan linfocitos,
clulas de la sangre mediante puncin o clulas
provenientes del lquido amnitico.
Cultivo de las clulas. Las clulas obtenidas se someten
al procedimiento de un medio lquido de cultivo de
tejidos, con la finalidad de obtener una poblacin
celular en proceso de mitosis.
Obtencin de clulas en metafase. Despus de
transcurridos unas 48 - 72 horas de cultivar las clulas;
se le aade al medio de cultivo sustancias que detenga
la mitosis, un alcaloide como la colchicina, con la
finalidad que todas aquellas clulas que estn en la
etapa de metafase ya no prosigan la divisin celular.
Lisis de las clulas. A continuacin se centrifuga el
medio de cultivo para obtener una concentracin de
clulas en las diversas fases de la mitosis, al
centrifugado se la aade un medio lquido hipotnico
que debilite la membrana celular.
Se toma una gota de la solucin y se deja caer sobre la
superficie de un portaobjetos, a una distancia de unos
10 a 15 centmetros. Esta accin provoca que las
clulas, ante el impacto, estallen y el contenido nuclear
Figura Biol. Cel. 41. Representacin esquemtica de un cromosoma
mostrando sus diferentes componentes estructurales.
La apariencia morfolgica de los cromosomas en metafase
es la siguiente: estn constituidos por dos brazos (cada uno
de ellos corresponde a cada cromtida) unidos por una
constriccin primaria denominada centrmero al cual se le
adosan protenas nucleares que conforman el cinetocoro,
zona del cromosoma a la que se adosan los microtbulos del
huso mittico.
Los cromosomas humanos son de tres tipos de acuerdo al
tamao de los brazos y a la disposicin del centrmero o
constriccin primaria:
Metacntricos, las partes constituyentes de los brazos son
casi del mismo tamao. El centrmero esta situado en la
parte central de cada brazo
Submetacntricos, el centrmero est localizado de tal
manera que las porciones superiores de los brazos son un
poco ms cortos que las porciones inferiores.
Acrocntricos la constriccin primaria se localiza casi en el
extremo superior de los brazos, las porciones superiores de
los brazos aparecen como pequeos apndices.
Figura Biol. Cel. 42. Diferentes tipos de cromosomas .
Cromatina sexual.
En los mamferos el sexo est determinado por la presencia
en cada clula somtica de los cromosomas denominados
sexuales. En los individuos de sexo femenino el par de
cromosomas sexuales est constituido por dos cromosomas
XX, heredados durante la fecundacin tanto del ovocito
como del espermatozoide. En los individuos de sexo
masculino, el par de cromosomas sexuales est representado
por los cromosomas XY, el cromosoma X es herencia del
ovocito y el cromosoma Y lo hereda del espermatozoide.
En los individuos de sexo femenino slo uno de los
cromosomas X es activo, en el sentido de expresar rdenes
para la sntesis protenica (transcripcin), el otro cromosoma
X permanece inactivo durante toda la vida. La inactividad
de uno de ellos se determina al azar, en las etapas muy
tempranas del desarrollo embriolgico.
En el ncleo de las clulas somticas en interfase de
individuos de sexo femenino, el cromosoma X inactivo est
representado por un conglomerado muy condensado de
heterocromatina a la cual se le denomina cromatina sexual
o cuerpo de Barr, en honor del investigador que la describi
por primera vez. Barr la observ en neuronas y clulas
epiteliales de la mejilla bucal. En neuronas aparece como
una condensacin intensamente basfila adosada a la
superficie del nuclolo y en las clulas epiteliales se observa
en forma de un grumo semicircular relacionado
estrechamente con la cubierta nuclear. Posteriormente la
cromatina sexual se descubri, en la forma de un pequeo
apndice esfrico, sostenido por una prolongacin de
cromatina de uno de los lbulos del ncleo de los
neutrfilos (leucocito de la sangre): En conjunto, la
cromatina sexual de los neutrfilos semeja a un palillo de
tambor, nombre con el cual tambin se le identifica en esas
clulas.
La observacin de la cromatina sexual: cuerpos de Barr,
en clulas epiteliales o los palillos de tambor en
neutrfilos de un frotis de sangre, en un porcentaje que
alcance aproximadamente un 5% a 7% de las clulas
examinadas, permite hacer un diagnostico preliminar del
sexo gentico de un individuo.
Las clulas humanas que albergan en su ncleo todo el
genoma (46 cromosomas o 23 pares) se denominan
diploides (2n) o euploides, Las clulas germinales,
ovocitos y espermatozoides que slo poseen 23
cromosomas, uno de los pares homlogos, se llaman
haploides (1n). Cuando una clula somtica posee dos
ncleos (dos genomas) se denomina tetraploides (4n), por
ejemplo algunas clulas hepticas o neuronas ganglionares;
si contiene ms de dos ncleos (mas de dos genomas) se le
conoce como poliploides, por ejemplo los megacariocitos y
los osteoclastos. Cuando una clula presenta un nmero
anormal de cromosomas que no sean mltiplos de 1n, se le
designa como aneuploides, Las personas que sufren del
sndrome de Down o el sndrome de Klinefelter, poseen en
el genoma de sus clulas somticas, 47 cromosomas o
aquellas que sufren del sndrome de Turner slo poseen 45
cromosomas.
08/08/03 19:20 horas
Figura Biol. Cel. 43. Diversos tipos de localizacin y formas de cromatina
sexual
cido ribonucleico.
El otro cido nucleico es el cido ribonucleico; tiene una
estructura similar al DNA, con las diferencias que consisten
en que es una sola cadena de nucletidos, la pentosa es la
ribosa y en vez de la base timina (T), la cadena posee el
aminocido uracilo (U).
En el ncleo, el DNA sirve como plantilla para generar una
cadena complementaria de RNA, proceso denominado
transcripcin.
Existen tres tipos de enzimas conocidas como polimerasas
de RNA que catalizan la sntesis de las tres clases de RNA:
a)
El RNA mensajero (RNAm) es catalizado por la
polimerasa RNA II.
La transcripcin contina hasta que la RNA polimerasa
abandona la hebra complementaria de DNA.
b) El RNA de transferencia (RNAt) se cataliza mediante la
polimerasa RNA III.
La transcripcin se inicia cuando la RNA polimerasa
reconoce en el triplete de DNA el codon de inicio AUG y
finaliza cuando la enzima reconoce un sitio de terminacin
de la cadena complementaria de los codones de terminacin
UAA, UAG o UGA.
c)
El RNA ribosomal (RNA r) por la polimerasa RNA I.
Esta enzima se localiza en el nuclolo.
Acido ribonucleico mensajero (RNAm). Es el encargado de
transportar el cdigo gentico desde el ncleo hacia el
citoplasma, donde sirve como plantilla para la sntesis de
una protena especfica. Cada RNAm es una copia
complementaria de la zona del DNA que integra un gen.
Esto significa que cada RNAm est formado por una
secuencia de codones que expresan aminocidos
especficos.
Su estructura qumica tiene un codon de inicio (AUG) y uno
o ms codones de trmino (UAA, UAG UGA)que indican
el final de la sntesis de la protena.
El RNAm transcrito en el ncleo se transporta, a travs de
los poros nucleares, al citoplasma en donde se traduce en
una protena especfica.
La transcripcin de los genes es catalizada por las enzimas
RNA polimerasas. Este proceso se inicia cuando la RNA
polimerasa se une a una secuencia de DNA denominada
DNA promotor situada cerca de la regin que se va a
transcribir. Como resultado de este inicio, las dos hebras de
la espiral doble de DNA se separan y se abren frente al
promotor. En esas condiciones la RNA polimerasa tiene
acceso a la hebra patrn de DNA que permitir el inicio de
la sntesis o transcripcin de la hebra complementaria del
RNA. Esta hebra ser idntica a la otra hebra del DNA que
no se transcribir, excepto que, en vez del poseer el
aminocido timina, ste ser sustituido por el aminocido
uracilo.
La hebra de RNA transcripta
y la hebra de DNA
codificadora poseen la misma orientacin direccional
conforme se van abriendo las dos hebras de DNA.
La unin entre la RNA polimerasa y el promotor se produce
mediante una serie de protenas denominadas factores de
transcripcin que, primero se unen al promotor y luego a la
RNA polimerasa. Las dos hebras de DNA se abren en un
lugar de aproximadamente 15 a 20 bases antes del sitio de
transcripcin de la molcula de RNA. Una vez iniciada la
transcripcin, la RNA polimerasa se desplaza en direccin
5-----3 de la hebra codificadora forzando la abertura de las
dos hebras de DNA. La porcin de la hebra ya transcripta de
RNA se separa inmediatamente de la hebra patrn y queda
suspendida en el nucleoplasma, en tanto que las dos hebras
de DNA se vuelven a unir.
La transcripcin solamente se produce en la eucromatina, en
las hebras de DNA que miden 10 nm de dimetro; es decir
que tiene lugar en la unidad de condensacin del DNA
conocida como nucleosoma. En la actualidad an no se sabe
con certeza la manera cmo las RNA polimerasas actan
para abrir la doble hlice de DNA y permitir que la hebra
patrn enrollada alrededor de los nucleosomas pueda
transcribir el RNA.
En el caso de la transcripcin de RNA mensajero, la
molcula transcripta es una molcula larga, de hebra nica,
llamada RNAm precursor o primario (pre RNAm). La
molcula de RNAm primario debe modificarse para que
mediante agregados o supresiones de secuencia de bases
disminuya su longitud, muchas veces hasta la dcima parte,
antes de ser exportada al citoplasma. El RNA primario est
formado por segmentos de secuencias de codificacin
(exones) y segmentos de secuencias no codificadoras
(intrones). Los intrones deben eliminarse y los exones se
deben empalmar de manera conjunta para integrar
finalmente la molcula de RNAm madura que debe ser
exportada al citoplasma.
09/08/03 18:00 horas
La molcula de RNAm primario est constituida por una
serie de secuencias de intrones y exones. Durante el recorte
se eliminan los intrones mediante la intervencin de
espliceosoma, complejo de recorte formado por varias
partculas pequeas de ribonucleoprotena nuclear (snRNP)
o snurps. Los diferentes tipos de snurps son capaces,
utilizando el apareamiento de bases de hallar los sitios 3 y
5correctos para la escisin o corte de los extremos y luego
propiciar la unin de los segmentos de los exones para
unirlos y formar la nueva molcula del RNAm maduro. Los
intrones desprendidos son degradados posteriormente.
El RNA mensajero transporta la informacin gentica
del ncleo al citoplasma para servir como un molde
o plantilla para la sntesis de protenas.
cido ribonucleico ribosomal (ARNr).
El RNAr se sintetiza en la regin fibrilar (componente
fibroso) del nuclolo por actividad de la polimerasa I. La
transcripcin primaria origina el preRNAr, una molcula de
gran tamao de 13,000 nucletidos, llamada RNAr 45S. Al
nuclolo se incorporan, provenientes del ncleo, molculas
pequeas de RNAr 5S y ribonucleoprotenas sintetizadas en
el citoplasma, las cuales se unen a la molcula principal y
forman una partcula de ribonucleoprotena (RNP)
gigantesca. En la parte granulosa del nuclolo, la RNP es
procesada por protenas ribosomales para formar finalmente
las dos subunidades: las pequeas de 18S y las grandes de
40S, las que se trasladan a travs de los poros nucleares
hacia el citoplasma, lugar donde se ensamblarn cuando se
requiera la sntesis de protenas. Las subunidades pequeas
tienen un valor de sedimentacin de 40S (18S de RNAr +
33 protenas asociadas) y las grandes poseen un valor de
sedimentacin de 60S (RNA de 28S, 5.8 S y 5S + 49
protenas).
El RNA ribosomal integra, en el ncleo, complejos
constituidos por protenas y enzimas para formar
ribosomas.
Acido ribonucleico de transferencia (ARNt).
El cido ribonucleico de transferencia es una molcula
pequea de RNA, transcripta del DNA mediante la actividad
de la polimerasa III. Esta constituida por 80 nucletidos
aproximadamente; la hebra formada se pliega sobre si
misma para adoptar la forma de hoja de trbol.
La hebra de RNAt tiene dos regiones importantes, una de
ellas es el anticodn, secuencia de nucletidos necesaria
para reconocer el codon que transporta el RNAm; la otra es
la regin transportadora del aminocido, que reside en el
extremo 3 de la molcula. El RNAt es aminocilado en el
ncleo y tambin en el citoplasma, esta accin le facilita la
actividad funcional que desarrolla en el citoplasma.
Desempea la funcin de acarrear tripletes de aminocidos
activados al complejo ribosoma RNAm en donde se
acoplarn para constituir la protena.
El RNA de transferencia selecciona y conduce los
aminocidos activados al complejo ribosoma-RNA
mensajero para que se produzca la sntesis protenica.
10/08/03 16:12 horas
Ribosomas. Son organelos no membranosos constituido
principalmente por RNAr asociado a protenas nucleares.
Miden 25 nm de dimetro y 25 nm de largo. Son los
encargados de la sntesis de protenas en el citoplasma.
Cada ribosoma esta integrado por dos subunidades, una
grande y una pequea. Ambas subunidades son transcriptas
en el nuclolo y exportadas por separado hacia el citosol.
Las dos unidades permanecen aisladas en el citoplasma,
solamente se ensamblan cuando un RNA mensajero las une
para iniciar la sntesis de protenas. Las subunidades
pequeas tienen un valor de sedimentacin de 40S (18S de
RNAr + 33 protenas asociadas) y las grandes poseen un
valor de sedimentacin de 60S (RNA de 28S, 5.8 S y 5S +
49 protenas).
La subunidad pequea tiene un sitio de unin para el RNA
mensajero, un sitio P para ligar el peptidil-cido nucleico de
transferencia (RNAt) y un sitio A para la unin de
aminoacil-RNAt.
La subunidad pequea de 40S esta dividida parcialmente en
una cabeza y una base por una hendidura central. La
subunidad grande de 60S, es generalmente ms redonda;
posee una muesca que se relaciona con la hendidura de la
subunidad pequea. Ellas forman un canal o tnel por el
cual se acomoda o desliza la hebra de RNAm, para la
traduccin y sntesis de la molcula peptdica.
Los ribosomas son los responsables de la basofilia
citoplasmtica.
Se disponen de dos maneras: a) fijos, adosados a la
superficie externa de las cisternas del retculo endoplsmico
rugoso o granular, nombre que se asigna al R.E por la
disposicin que adoptan los ribosomas con relacin a sus
cisternas y b) ribosomas libres en el citoplasma. La gran
mayora de clulas que existen en el cuerpo humano, con
excepcin de los eritrocitos y clulas del estrato crneo de la
epidermis, poseen ribosomas libres en menor o mayor
cantidad.
Figura Biol. Cel. 44. Imagen de un ribosoma: se observan las unidades
mayor y menor. Estructura de un polirribosoma.
Los ribosomas fijos y los libres se renen en cadenas,
denominadas polirribosomas integrados por unos pocos
hasta ms de 30 unidades, as agrupados forman rosetas,
espirales o semicrculos. La unin de los ribosomas se
produce por la presencia de una hebra de ARN mensajero
(RNAm) que mide aproximadamente de 1 a 1.5 nm de
longitud.
Los ribosomas adheridos al RER se encargan de sintetizar
protenas de exportacin, aquellas que constituyen la
secrecin de las clulas; protenas luminales, que se
integran en forma de protenas membranales de organelos
como el retculo endoplsmico, las cisternas del aparato de
Golgi, los lisosomas y otras vesculas citoplasmticas y la
sntesis de protenas integrales del plasmalema.
Sntesis de protenas. Se produce en los ribosomas
adosados al RER y en los ribosomas libres suspendidos en
el citoplasma.
El procedimiento para sintetizar una protena requiere de los
componentes celulares siguientes:
Una hebra de RNA mensajero.
Subunidades ribosomales grandes y pequeas
Varias molculas de RNA de transferencia; cada una de
ellas transporta un aminocido y posee el anticodn que
reconoce el codn transcripto al RNAm responsable de
codificar el aminocido especfico.
Sntesis de protenas en el citoplasma. Las protenas que
no van a salir de la clula se sintetizan en los polirribosomas
libres; as tenemos que las protenas lisosomales, protenas
membranales de organelos citoplasmticos, protenas
perifricas de la superficie interna o citoslica del
plasmalema cuyos enlaces son dbiles como la espectrina y
la anquirina; todas aquellas protenas que formarn parte
del citoesqueleto y protenas funcionales, como la
hemoglobina, actina y miosina y las nucleoprotenas. La
sntesis de estas protenas se produce mediante las etapas
siguientes:
El procedimiento se inicia cuando el sitio P de la
subunidad pequea es ocupado por un RNAt iniciador,
cuyo anticodn reconoce al codn AUG que expresa el
aminocido metionina.
A la subunidad pequea se le adosa un RNA mensajero.
La subunidad pequea colabora con el anticodn de la
molcula de RNAt a reconocer el codn de inicio AUG
en la hebra de RNAm. Esta etapa interviene como una
fase de registro, as se pueden reconocer los tres
nucletidos siguientes de la hebra de RNAm como el
codn inmediato.
La subunidad grande se liga a la subunidad pequea y el
ribosoma se desliza a lo largo de la hebra de RNAm en
direccin de 5 a 3, hasta que el codn siguiente se
alinea con el sitio A de la subunidad pequea.
Un RNAt acilado (que transporta un aminocido
compara su anticodn con el codn del RNAm; si
corresponden, el RNAt se une al sitio A.
Los aminocidos forman en los sitios A y P un enlace
peptdico.
En el sitio P el RNAt cede su aminocido al RNAt
localizado en el sitio A que exhibe ahora dos
aminocidos unidos a l. La enzima transferasa de
peptidilo cataliza estas reacciones.
El primer RNAt sin su aminocido abandona el sitio P;
el segundo RNAt con dos aminocidos unidos se
traslada del sitio A al sitio P. Esto produce que el
ribosoma se mueva a lo largo de la hebra de RNAm
hasta que se alinea el siguiente codn con el sitio A de la
subunidad pequea. La energa necesaria para este
movimiento se genera por hidrlisis de GTP.
Estas etapas se repiten, alargndose la cadena de
polipptidos hasta que el ribosoma llega al codn de
terminacin o detencin. Los tres codones de
terminacin son UAG, UAA y UGA; cualquiera de ellos
puede detener la traduccin.
Cuando en el sitio A de la subunidad pequea llega un
codn de terminacin, se liga al sitio A un factor de
liberacin que tiene como funcin liberar la cadena
polipeptdica (protena primaria) recin sintetizada del
sitio P al citoplasma.
El ltimo RNAt se libera del sitio P y tambin el factor
de liberacin del sitio A, en tanto que las subunidades
ribosomales pequea y grande se desligan del RNAm.
Figura Biol. Cel. 45. Representacin esquemtica de la sntesis de
protenas por los polirribosomas citoplasmticos.
Mecanismo de la sntesis de protenas en el retculo
endoplsmico rugoso.
Aquellas protenas que deben ser exportadas de la clula en
la forma de secrecin o ser aisladas del citosol para integrar
componentes protenicos de las membranas celulares, deben
sintetizarse e incorporarse a la cisterna del RER.
La identificacin del procedimiento se inicia con un
segmento pequeo de RNAm localizado despus del codn
de inicio, que codifica una cadena pequea de aminocidos
llamada pptido de seal. Este es reconocido como tal por
un complejo protena-RNA, existente en el citoplasma, la
partcula de reconocimiento de seal (PRS) Esta partcula
unida al pptido de seal, localizada en el sitio P de la
subunidad pequea, detiene la traduccin y gua el RNAr
unido al RNAm (polirribosoma) al RER.
protenicas delgadas que se entrecruzan a travs del espacio
nuclear. Este aspecto se logra observar despus de aplicar
mtodos de extraccin empleando detergentes no inicos y
soluciones con altas concentraciones de sal que eliminan
lpidos, y la gran mayora de protenas histonas y no
histonas de la cromatina. Posteriormente se procede a
digerir las hebras de DNA con la enzima
desoxiribonucleasa, permaneciendo, al final, la fina red
fibrilar mencionada.
Figura Biol. Cel. 46. Representacin esquemtica de la sntesis de
protenas por los polirribosomas adheridoa al RER.
La protena receptora del PRS, llamada protena de anclaje o
de desembarque localizada en el citoplasma, se une con el
PRS; en tanto la partcula grande del ribosoma se une a la
protena citoslica transmembranal receptora, de esa manera
el complejo polirribosomal est ensamblado para iniciar la
sntesis de la protena especfica. Para alcanzar ese objetivo
deben producirse los procesos siguientes:
Ensamble de protenas integrales del RER para formar
un poro que atraviese la doble capa de lpidos.
El pptido seal se pone en contacto con el poro
protenico y empieza a introducirse, primero la terminal
amino, hacia la cisterna del RER.
La partcula PRS se desprende del sitio P de la partcula
pequea del ribosoma y vuelve al citoplasma. El
ribosoma permanece en contacto con el RER.
A medida que se reanuda la traduccin, la protena que
se forma, conectada al pptido seal, se introduce a la
cisterna del RER.
Una enzima localizada en la superficie cisternal del
RER, llamada peptidasa de seal, corta el pptido de
seal de la protena que se est formando. El pptido
seal por accin enzimtica se degrada en sus
componentes de aminocidos.
Cuando la traduccin llega al codn de detencin, la
sntesis de la protena termina, las partculas pequeas y
grandes se disocian y vuelven al citosol y formar la
reserva de subunidades ribosomales.
Las protenas recin formadas, se pliegan, glucosilan y
experimentan modificaciones postraduccionales dentro
de las cisternas del RER.
Las protenas sintetizadas abandonan el RER dentro de
pequeas vesculas que se dirigen a porciones del RE
carentes de ribosomas (RE transicional) y de all al
RECIG.
11/08/03 12:40 horas
Matriz nuclear.
Se define a la matriz nuclear como aquella estructura
constituida por una red fibrilar insoluble que adopta la
forma del ncleo, la integran un conjunto de fibrillas
La matriz nuclear sirve como un esqueleto o armazn que
mantiene la forma del ncleo o es un molde sobre el cual se
organizan las asas de cromatina. Tambin permite el anclaje
del mecanismo comprometido en las diferentes actividades
del ncleo tales como la transcripcin, duplicacin del
RNA. Experimentos realizados para tal efecto demuestran
que el RNA transcripto permanece en contacto con el gen
del DNA que lo gener, sin apartarse del mismo, esto indica
que la trama fibrilar fina de la matriz nuclear le impide
alejarse del lugar donde se origin.
Nuclolo.
Es una estructura ovalada o esfrica, densa, no membranosa,
localizada en el interior del ncleo. Dependiendo el
momento funcional de la clula y su capacidad para la
sntesis de protenas suelen existir uno, dos o tres nuclolos.
El nuclolo nicamente se observa en la interfase de la
clula; mide aproximadamente un micrmetro de dimetro,
aunque en las neuronas de la sustancia gris de la mdula
espinal, en los hepatocitos y en las clulas glandulares de
secrecin rica en protenas pueden medir hasta 2
micrmetros de grosor.
Figura Biol. Cel. 47. Fotomicrografias y esquema del nucleolo observado
mediante a) microscopio fotnico y B) microscopio electrnico de
transmisin. c) esquema del nucleolo.
Durante la mitosis no es posible distinguirlo porque sus
componentes se disgregan. Con la tincin de hematoxilina
eosina muestra una reaccin basfila, aunque a veces
adopta una coloracin ligeramente acidfila (debido a la
presencia de abundantes ribonucleoprotenas) Posee escasa
cantidad de DNA inactivo por lo tanto no se tie con el
reactivo de Feulgen. Las regiones nucleolares que se tien
intensamente (la cromatina asociada al nucleolo)
corresponden a la cromatina que se asocia ntimamente con
el nucleolo, encargada de transcribir RNA ribosomal.
licuadas se rompan y liberaban la enzima. El empleo de
tcnicas histoqumicas para fosfatasa cida facilit la
demostracin de pequeas partculas que reaccionaban
positivamente a la determinacin de la enzima y que
pertenecan a la fraccin mitocondrial.
En las clulas donde existe una gran actividad de sntesis de
protenas el nucleolo puede alcanzar un volumen
equivalente al 20 - 25 % del total del ncleo.
El microscopio electrnico muestra cuatro reas diferentes
constitutivas del nucleolo:
Componente granular. Es el lugar donde se ensamblan
subunidades ribosomales en maduracin. Est formada
por grnulos electrndensos de ribonucleoprotena que
miden unos 15 nm de dimetro.
Componente fibrilar denso. Constituido por RNA
nucleolares en proceso de transcripcin.
Centros fibrilares de tincin plida. Formados por
hebras de DNA inactivo (no transcribe RNAr). En estas
regiones del nucleolo, en clulas de la especie humana,
se localizan los extremos de los cromosomas 13, 14, 15,
21 y 22 que contienen las regiones de organizacin
nucleolar (RON), en donde se localizan los locus de
genes que codifican el RNA ribosomal.
Matriz nucleolar. Red fibrilar fina que interviene en la
organizacin y soporte de los componentes del
nuclolo.
Lisosomas.
La existencia de los lisosomas fue propuesta en la dcada de
los aos 50, por Christian de Duve, investigador belga que
posteriormente fue galardoneado con el premio Nobel,
gracias a la serie de experimentos que realiz fue posible
demostrar la existencia de los lisosomas.
Los experimentos se disearon para investigar la presencia
de las enzimas hepticas, glucosa 6 fosfatasa y fosfatasa
cida. Para tal fin se utilizaron homogenizados de hgado de
los cuales mediante centrifugacin diferencial, se obtuvieron
fracciones mitocondriales, nucleares, microsomal y citosol.
En esas fracciones se logr demostrar que la enzima glucosa
6 fosfatasa se encontraba presente en la fraccin
microsomal, constituida principalmente por micropartculas.
La determinacin de la fosfatasa cida solamente fue
posible hacerla cuando el homogenizado de hgado era
sometido a fraccionamiento por una licuadora de alta
velocidad, pues en el anterior experimento la cantidad de
fosfatasa cida era mnima, en cambio cuando este
homogenizado se disgregaba en forma drstica y luego se
congelaba, se poda detectar una cantidad considerable de la
enzima presente en la fraccin mitocondrial.
Este hecho le permiti a de Duve, inferir que la fosfatasa
cida se localizaba dentro de las mitocondrias porque al ser
Figura Biol. Cel. 48. Lisosomas observados como a) en leucocitos
neutrfilos y b) al microscopio electrnico y ptico
El microscopio electrnico demostr que esas pequeas
partculas eran vesculas membranales que albergaban en su
interior un contenido homogneo o ligeramente granular, de
morfologa diferente a la exhibida por las mitocondrias. Esta
primera demostracin se confirm an ms cuando se
demostraron dentro de estas vesculas otras hidrolasas
cidas como la ribonucleasa cida, la desoxiribonucleasa
cida, la betaglucoronidasa y la catepsina.
La presencia de varias enzimas, contenidas en una misma
vescula, que actuaban de forma ptima, en un medio de pH
cido y sobre diferentes substratos con resultados de efecto
ltico, motiv que de Duve les denominara lisosomas.
Desde esa poca hasta la actualidad se han logrado
demostrar una gran cantidad de enzimas en el interior de los
lisosomas, cada una de ellas acta siempre en un medio
cido. La mayora de las sustancias qumicas presentes en
las clulas tales como protenas, carbohidratos, lpidos y
cidos nucleicos son degradadas por las enzimas
lisosomales. La existencia de la membrana que las
constituye evita que estas enzimas se dispersen en el
citoplasma. Varias actividades fsicas y qumicas tales como
el fro, la radiacin ultravioleta, los ultrasonidos, los
disolventes orgnicos o la disminucin del aporte de
oxgeno, falta de nutrimentos, suelen provocar la rotura
parcial o total de la membrana y el vaciamiento de las
enzimas y la destruccin de la clula.
Esto mismo suele suceder en procesos patolgicos, lo que
causan las modificaciones que sufren las clulas y los
tejidos despus de la muerte celular, los cambios
ocasionados en las clulas se denominan en conjunto,
autlisis o degeneracin postmortem. Esta forma de muerte
celular, no solamente es patolgica, tambin sucede en
clulas
embrionarias
en
condiciones
normales,
especialmente cuando ciertas clulas deben morir para
permitir la proliferacin de otras poblaciones celulares o
iniciar procesos de diferenciacin tisular. Procesos
infecciosos o traumticos tambin provocan la actividad
lisosomal para destruir las clulas daadas.
Debe tenerse en cuenta que de Duve demostr la existencia
de los lisosomas en base nicamente a fundamentos
bioqumicos. Fue Novikoff, en 1955, junto con de Duve, al
examinar fracciones obtenidas mediante centrifugacin
fraccionada de hepatocitos, utilizando el microscopio
electrnico, constataron una intensa
actividad de la
fosfatasa cida en componentes microsomales, cuya
apariencia era la de pequeas vesculas membranales de
caractersticas morfolgicas totalmente diferentes a las
mitocondrias.
Formas de lisosomas.
Con el microscopio electrnico se han logrado identificar
varios tipos de lisosomas. Las diversas formas que los
lisosomas poseen reflejan las diferencias del estado
funcional en determinados momentos por los que pasan las
clulas.
La digestin de sustancias o partculas extraas
incorporadas al interior de las clulas (endocitosis: picnosis
o fagocitosis) o la digestin de componentes celulares que
deben destruirse por haber terminado su ciclo funcional
(autofagia o endofagocitosis), son los momentos en el ciclo
vital de las clulas donde los lisosomas muestran formas
variables y su actividad digestiva.
A continuacin se muestra en un cuadro varias enzimas
lisosomales y los substratos correspondientes:
Enzimas lisosomales
Catepsinas A, B, C, D y
E
Colagenasa
Peptidasa
Ribonucleasa cida
Desoxirribonucleasa
cida
Fosfatasa cida
Beta-galactosidasa
Alfa-glucosidasa
Beta-glucosidasa
Sialidasa
Lisozima
Hialuronidasa
Fosfolipasa
Esterasa
Substratos
Varios tipos de protenas.
Colgena
Varios pptidos
Acido ribonucleico
Acido desoxirribonucleico
Monosteres de fosfato
Beta-galactsidos
Glucgeno
Beta-glucsidos
cido silico.
Pared
bacteriana,
mucopolisacridos
Acido
hialurnico,
condroitinsulfato.
Lecitina, fosfatidiletanolamina
steres de cidos grasos
12/08/03 20:25 horas
Los lisosomas son un conjunto de organelos membranosos
que varan de manera notable en forma y tamao. No es
fcil identificarlos por su tamao nicamente. Tanto el
microscopio fotnico y el electrnico muestran imgenes de
estructuras granulares o vesiculares, respectivamente que no
son fciles de identificar. La variedad en el tamao, abarca
dimensiones que oscilan entre 0.1 a 0.8 micrmetros, estn
limitados por una sola membrana y el contenido es poco
electrondenso. La mejor manera de identificarlos es
basndose en la aplicacin de mtodos citoqumicos, para
demostrar el contenido de una variedad de enzimas propias
de los lisosomas. Uno de ellos, mejor conocido es el mtodo
de Gomori, desarrollado en 1952, que identifica la presencia
de fosfatasa cida. Las imgenes de esta reaccin pueden
observarse utilizando los microscopios fotnico y
electrnico.
Figura Biol. Cel. 49. Fotomicrografas electrnicas de diferentes formas y
tamaos de lisosomas. Fotomicrografa electrnica de una mitocondria
siendo digerida por la fusin de un lisosoma para constituir un
autofagosoma .
Los lisosomas se originan por gemacin, desde el RTG del
aparato de Golgi, en forma de pequeas vesculas que
contienen las hidrolasas cidas. Las enzimas se sintetizan en
el RER en la forma de glucoprotenas (manosa + protenas
especficas). La manosa se fosforila en el aparato de Golgi
transformndose en manosa-6-fosfato que interviene como
un marcador especfico para sealar el contenido como
perteneciente a un lisosoma.. El reticulado trans de Golgi
contiene protenas integrales en su membrana que actan
como receptores de manosa-6-fosfato orientados hacia la luz
de las cisternas. En este lugar las enzimas lisosomales se
ligan a los receptores de manosa-6-fosfato; a continuacin
se produce la liberacin de vesculas recubiertas por
clatrina. Liberadas las vesculas, inmediatamente despus se
separa la cubierta de clatrina. A estas vesculas que
contienen las enzimas lisosomales se les denominan
lisosomas primarios. Algunos lisosomas primarios pueden
llegar a la membrana celular y mediante endocitosis, liberar
las enzimas lisosomales, como ocurre en los osteoclastos,
clulas responsables de la resorcin del tejido seo.
Otros tipos de lisosomas son los lisosomas secundarios, se
designan as a las vesculas que contienen hidrolasas cidas
y material que debe ser degradado. De acuerdo al tipo de
material que debe ser digerido, los lisosomas secundarios se
clasifican como: fagolisosomas o heterolisosomas y los
autofagosomas o autolisosomas. Los heterolisosomas se
forman por la fusin de lisosomas primarios con vacuolas
citoplasmticas que contienen sustancias extracelulares
transportadas al interior de la clula por algunos de los
procesos de endocitosis: fagocitosis y pinocitosis. Cuando
se produce la fusin del heterolisosoma con la vacuola
endoctica, las enzimas hidrolticas se vierten al interior de
la vescula originando un fagosoma. Los autolisosomas se
forman por la fusin de lisosomas primarios con
componentes
citoplasmticos
como
mitocondrias,
fragmentos de retculo endoplsmico rugoso o liso,
microcuerpos etc. La autodigestin de organelos celulares es
un acontecimiento normal en la vida de las clulas
especialmente durante la diferenciacin, crecimiento y
reparacin celular. Conforme transcurre l a digestin las
enzimas degradan las sustancias en proceso de degradacin,
Existen dos vas de los productos de la digestin, ciertos
compuestos tiles (aminocidos, azucares, etc.) para la
clula son incorporados al citosol, a travs de la membrana
mediante difusin pasiva o de transporte activo, para ser
utilizados posteriormente. Otros productos no digeribles se
eliminan o se excretan hacia el espacio extracelular
mediante exocitosis.
Figura Biol. Cel. 50. Lisosomas con sustancias no digeridas en su interior::
a) pigmento de lipofucsina en fibras musculares cardiacas, b) pigmento de
lipofucsina en neuronas ganglionares y c) fotomicrografa electrnica de
un cuerpo residual.
Cuando las sustancias endocitadas y organelos
autofagocitados no son digeridos totalmente, los residuos
permanecen dentro del compartimento membranal lisosomal
y se transforma en un cuerpo residual, conocido
antiguamente como grnulos o pigmentos de lipofucsina,
frecuentes en el citoplasma de las clulas que tienen una
vida muy larga (neuronas y fibras musculares cardiacas).
Los cuerpos residuales permanecen por mucho tiempo en el
interior de las clulas, donde se van acumulando en nmero
y tamao, esto sucede en clulas que por su larga vida
comienzan a envejecer. Al pigmento de lipofucsina
(acumulacin de los cuerpos residuales) se le denomina
tambin como pigmento de envejecimiento o de
desgaste.
13/08/03 23:30 horas
Los lisosomas y otras vesculas generadas durante el
proceso de endocitosis, poseen en sus membranas una
bomba de protones activada por la presencia de ATP, que se
encarga de impulsar iones de hidrgeno al interior de las
vesculas con la finalidad de acidificar el contenido hasta
alcanzar un pH de 5, ptimo para la actividad de las enzimas
lisosomales. A causa de la disminucin del pH, las enzimas
rompen su unin con los receptores de manosa-6-fosfato, se
desprenden de la membrana y se solubilizan en la luz de las
vesculas, lugar en donde una fosfatasa lisosmica separa el
fosfato de la manosa e impide una nueva unin con los
receptores. Estos ltimos se integran nuevamente cuando las
vesculas se reciclan al aparato de Golgi, all se fusionan
con las membranas con el propsito de ser reutilizados
nuevamente.
Funciones de los lisosomas:
Lisis de organelos durante el crecimiento, proliferacin y
diferenciacin celular.
Destruccin de microorganismos (protozoarios, virus y
bacterias) y partculas extraas al organismo por accin
de los macrfagos.
Eliminacin de partes celulares envejecidas y protenas
desnaturalizadas.
Proceso de queratinizacin de epitelios (epidermis, por
ejemplo).
Destruccin de eritrocitos viejos y clulas muertas.
Liberacin
de
hidrolasas
del
acrosoma
de
espermatozoides para que penetren en el ovocito,
durante la fecundacin.
Digestin de vitelo durante el desarrollo embrionario.
Resorcin del hueso.
14/08/03 14:00 horas
Endocitosis.
Se denomina endocitosis a la funcin mediante la cual la
clula incorpora desde el microambiente extracelular
sustancias a su economa, en la forma de partculas,
macromolculas y lquidos que no atraviesan la membrana
celular utilizando el transporte pasivo o el activo.
El material endocitado es incorporado al interior del citosol
mediante dos procedimientos:
A) Fagocitosis. Se conoce con este nombre al proceso
mediante el cual la partcula o macromolcula es
englobada en una vescula grande que por lo menos
mide ms de 250nm de dimetro; por ejemplo, bacterias,
restos celulares, eritrocitos viejos o muertos.
Las clulas responsables de realizar esta funcin se
denominan fagocitos. Los neutrfilos y los monocitos
(tipos diferentes de leucocitos o glbulos blancos) son
los fagocitos ms comunes en un organismo animal.
Ambas clulas para ejercer la funcin de fagocitosis
abandonan el torrente sanguneo y en el tejido
extravascular desarrollan eficazmente esta capacidad. En
estas circunstancias los monocitos adoptan el nombre de
macrfagos. Los fagocitos pueden captar partculas de
diversa naturaleza porque en su plasmalema poseen
receptores que identifican al material y lo fagocitan
mediante evaginaciones de la membrana en la forma de
finos seudpodos o filopodios que rodean a la partcula,
la rodean o envuelven y de esa manera es transportada,
envuelta en una vescula membranal, al interior del
citoplasma. La vescula que contiene el material
fagocitado se denomina fagosoma.
fotnico. Es posible afirmar que todas las clulas del
organismo realizan este tipo de endocitosis.
Macropinocitosis. La clula tambin capta sustancias sin
seleccionar que se incorporan en vesculas de mayor
tamao, visibles con el microscopio fotnico (pueden
medir varios micrmetros de dimetro). Las clulas del
tbulo contorneado proximal del rin son un ejemplo
de este transporte. En el tercio apical de estas clulas se
observan una gran cantidad de vesculas pinocticas;
ellas son las responsables de devolver al torrente
circulatorio una gran cantidad del plasma filtrado para
formar la orina (En la especie humana los riones filtran
aproximadamente 200 litros de sangre en 24 horas y
deben reabsorberse el 99% para eliminarse como orina
solamente el 1%).
Figura Biol. Cel. 51. Esquema que representa las diversas fases de la
fagocitosis.
B) Pinocitosis. Es la captacin de porciones pequeas de
lquidos en las que se encuentran disueltas o en
suspensin partculas de bajo peso molecular. La gran
mayora de clulas eucariotas efectan continuamente
pinocitosis. Es la forma de regular de forma eficiente el
equilibrio hdrico entre el ambiente extracelular y el
contenido citoplasmtico. Tambin le sirve a la clula
para mantener constante la cantidad de componentes del
plasmalema, por cuanto tambin se produce una
permanente exocitosis, especialmente en clulas cuya
funcin preponderante es la secrecin ecrina o
mercrina. El equilibrio en la cantidad de material de la
membrana consiste en que, a travs de la pinocitosis y la
fagocitosis el plasmalema pierde material al formar
vesculas grandes o pequeas en tanto la membrana
aumenta en componentes cuando por exocitosis los
grnulos de secrecin o las vesculas que contienen
residuos o desechos del catabolismo celular se unen y
fusionan al plasmalema para excretarlos.
Existen tres maneras por las cuales una clula ejerza la
pinocitosis. Cada tipo de pinocitosis depende del tamao
de las vesculas formadas, la seleccin del material que
se endocita y el mecanismo por el cual se realiza la
captacin de sustancias. DE acuerdo a lo expuesto la
pinocitosis puede ser:
Micropinocitosis. Es la incorporacin no selectiva de
lquidos con molculas disueltas que se efecta mediante
pequeas vesculas, no visibles con el microscopio
Figura Biol. Cel. 52. Representacin esquemtica de la s diversas fases de
la pinocitosis.
Pinocitosis mediada por receptor. Es una forma de
pinocitosis en la que determinadas macromolculas se
incorporan a la clula en forma selectiva, por
reconocimiento especfico de receptores membranales.
De esta manera se captan hormonas, y factores de
crecimiento, que mediante ligandos se unen con los
receptores. Al unirse el ligando con el receptor, se
induce a una leve invaginacin de la membrana,
causada por la adhesin, en la superficie interna del
plasmalema de molculas de la protena clatrina; las
pequeas depresiones se
denominan fositas
recubiertas. Poco tiempo despus las fositas se
invaginan an ms y se transforman en vesculas
recubiertas, diferencindose de las otras vesculas
pinocticas, porque estas ltimas tienen lisa la superficie
citoslica de la membrana.
Digestin celular:
Cualquiera que sea la modalidad de endocitosis (fagocitosis
o pinocitosis), la clula incorpora sustancias slidas o
lquidas siempre rodeadas de vesculas membranosas,
cubiertas o no de molculas protenicas.
Todas las vesculas endocticas reciben el nombre de
fagosomas. En los fagosomas con cubierta de clatrina, las
partculas a ser endocitadas se unen a receptores
membranales ubicados en la superficie extracelular del
plasmalema; de manera simultnea los trisquiliones se
adosan a la superficie interna de la membrana celular para
formar la fosita cubierta de clatrina y constituir despus, la
vescula de clatrina que se desprende del plasmalema para
transformarse en un fagosoma cubierto. Posteriormente los
trisquiliones de clatrina se desacoplan de la vescula y son
reciclados al citoplasma en las cercanas de la superficie
interna de la membrana celular. En esas condiciones el
material endocitado queda dentro de una vescula
descubierta la cual vaca su contenido en un endosoma
temprano para constituir un endosoma tardo o lisosoma
maduro. La vescula vaca de la partcula endocitada se
traslada al plasmalema para el reciclamiento de los
receptores membranales.
Por otro lado, del RTG se forman vesculas conteniendo
enzimas lisosomales inactivas con un pH de 6.0,
denominadas por algunos autores como prelisosomas o
endosomas tempranos, marcados con manosa-6-fosfato,
los cuales se cubren con trisquiliones de clatrina, en ese
momento la bomba de hidrgeno incorpora al endosoma, de
manera activa, iones de H+ acidificando el interior a un pH
de 5.5. El ambiente cido genera desacoplamiento del
ligando desde su molcula receptora. Las molculas
receptoras se liberan y vuelven hacia el plasmalema, en
tanto que el ligando permanece dentro de la vescula. Si la
partcula que contiene el fagosoma debe ser digerida en su
totalidad, el endosoma temprano se desliga de la cubierta de
clatrina y se pone en contacto con el fagosoma, fusionan
sus membranas y el contenido del endosoma temprano se
vierte en el fagosoma. Al complejo formado endosoma
temprano + fagosoma se le conoce como endosoma tardo,
o lisosomas maduros.
Las enzimas hidrolticas digieren la mayora de los
componentes del contenido del fagosoma especialmente
carbohidratos y protenas y los transforman en sustancias
solubles que son transferidos al citosol, mediante protenas
transportadoras, los productos son reutilizados por la clula.
El resto del material endocitado que no pudo ser digerido, es
devuelto al espacio extracelular mediante exocitosis como
sustancia de desecho.
Los lpidos son ms resistentes a la digestin completa y se
quedan encerrados dentro del lisosoma, transformando al
lisosoma gastado o agotado (en su funcin enzimtica) en
un cuerpo residual; conocido mediante microscopa
fotnica, como grnulo o pigmento de lipofucsina o
pigmento de desgaste o envejecimiento.
Peroxisomas.
Dentro de la fraccin microsomal del citosol, existen
pequeos
organelos
membranosos
denominados
peroxisomas de forma esfrica u ovoidea que miden 0.2 a
0.5 micrmetros de dimetro. Contienen ms de 40 enzimas
oxidativas del tipo catalasa, uratooxidasa y aminooxidasa,
acil-CoA oxidasa, etc. Existen en la gran mayora de las
clulas animales, especialmente en clulas del hgado (de
Duve, en 1965 demostr que los microcuerpos del hgado de
rata contenan oxidasas que transferan tomos de hidrgeno
al oxgeno molecular formando perxido de hidrgeno) y
de los riones. Tambin se han observado en clulas
vegetales. El contenido de los peroxisomas muestra un
aspecto homogneo, finamente granular. En algunas
especies animales y en ciertas clulas vegetales (hojas del
tabaco) los peroxisomas poseen una estructura cristaloide
electrondensa denominada nucleoide constituida por la
enzima uratooxidasa.
La cantidad y variedad de enzimas son propias de cada tipo
celular. La enzima catalasa es la nica que existe en todos
los peroxisomas de las clulas.
Figura Biol. Cel. 53. fotomicrografa electrnica de un peroxisoma.
Las enzimas de los peroxisomas desarrollan varias
funciones; tienen la capacidad de desintoxicar sustancias
dainas para el organismo, as tenemos que actan contra el
etanol, metanol, fenoles y formaldehido, de all la presencia
copiosa de ellos en las clulas hepticas y renales.
Degradan los cidos grasos de cadena larga (oxidacin beta)
para formar acetil CoA y H2O2 ( perxido de hidrgeno o
agua oxigenada) pues combina el hidrgeno del cido graso
con oxgeno molecular (O2 ). La acetilCoA es utilizada por
las clulas para su propio metabolismo o la exporta hacia el
espacio extracelular para que la usen clulas vecinas. El
perxido de hidrgeno mata a microorganismos. La catalasa
degrada al H2 O2 que no se utiliza.
Se ha constatado que los peroxisomas se reproducen de
otros peroxisomas como las mitocondrias. Las protenas
membranales se originan en polirribosomas libres o
citoslicos, de all se trasladan a las paredes citoplasmticas
de los peroxisomas.
Mitocondrias.
Las clulas animales y vegetales necesitan de energa para
realizar sus funciones. La energa se genera por
transformacin de la adenosina trifosfato (ATP), sustancia
rica en energa. Para liberar la energa endergnica que
posee debe transformarse en adenosina difosfato (ADP) o
en ciertos casos en adenosina monofosfato (AMP). En las
clulas existen tres vas para producir esta energa qumica:
El ATP se sintetiza en el citoplasma durante la cadena
de reacciones exergnicas denominada gluclisis,
proceso en el cual los carbohidratos son catabolizados.
El ATP se genera dentro de los organelos llamados
mitocondrias, existentes en todas las clulas eucariotas.
El ATP se forma en la mayora de las clulas vegetales,
dentro de unos organelos conocidos como cloroplastos.
En la clula animal las mitocondrias son las responsables de
generar la energa funcional, aunque estos organelos
tambin desarrollan otras funciones vitales para la clula.
Klliquer, en la dcada de 1870 describi unas partculas en
el msculo estriado de insectos y estudi su comportamiento
osmtico en varias soluciones salinas. Dedujo que eran
estructuras independientes no asociadas directamente con
los otros componentes celulares. En 1890 Altmann,
utilizando una coloracin constituida principalmente por
fucsina cida, describi grupos de estos grnulos en el
interior de clulas que aparentemente desarrollaban una gran
utilizacin de energa, los denomin bioblastos; trmino
que fue reemplazado por Benda por el mitocondria ( del
griego, mito= filamento o hilo + chondrion = grnulo) con
relacin a las formas filamentosas o granulosas de estas
partculas, cuando se observan al microscopio fotnico. En
1900, Michaelis utiliz el colorante vital verde Jano en
clulas vivas para colorear, selectivamente, a las
mitocondrias. Las reacciones oxidativas de las mitocondrias
producen el cambio de color del verde Jano, previamente
decolorado, a su color original de tal manera que las
mitocondrias se tien intensamente de un color verde,
pudindolas identificar a plenitud, de manera especfica. El
verde Jano se emplea como marcador citoqumico de las
mitocondrias en clulas vivas.
En 1930 Claude, Bensley y Hoerr, lograron aislar de
homogenizados celulares, fracciones puras de mitocondrias,
en ellas demostraron que eran los componentes celulares
donde se producan las reacciones oxidativas. De manera
casi simultnea Krebs descubri las diversas reacciones del
ciclo tricarboxilico generador de energa (ciclo de Krebs),
En los aos de la dcada de 1950, Lehninger y otros
colaboradores confirmaron que las mitocondrias realizaban
las funciones antes citadas y tambin las de la oxidacin de
los cidos grasos y la fosforilacin oxidativa (gnesis de
ATP).
Figura Biol. Cel. 54. Fotomicrografias de mitocondrias observadas
mediante del microscopio fotnico.
Estructura de la mitocondria. Las mitocondrias son
organelos lo suficientemente grandes para que, con
tinciones especiales, puedan ser observadas con el
microscopio fotnico. Poseen caractersticas morfolgicas
que permiten distinguirlas de otros organelos, pueden ser
esfricas, en forma de bastones o filamentosas. Suelen
medir entre 0,5 a 10 micrmetros de longitud o a veces unos
micrmetros ms. La gran variabilidad de formas y tamaos
dependen del tipo y funciones de las clulas que las
contienen.
Tambin existen grandes variaciones en cuanto a la
cantidad de ellas. As, un hepatocito suele poseer unas 1,500
a 2,500 mitocondrias, casi el 20% de su volumen total. Lo
mismo se observa en las clulas (fibras) musculares
esquelticas y cardiacas, clulas en las cuales existe una
gran demanda de energa para generar contraccin muscular.
Tambin se observa abundancia de mitocondrias en las
clulas parietales del estmago (son las clulas que
sintetizan el cido clorhdrico (HCl), en la pieza intermedia
de los espermatozoides (energa necesaria para producir el
movimiento flagelar), en tercio apical de las clulas
intestinales ( incorporacin de sustancias al interior de las
clulas mediante transporte activo) o en el tercio inferior de
las clulas del tbulo contorneado renal (devolucin al
torrente sanguneo de casi el 80% del liquido filtrado por los
corpsculos renales.
Mediante el empleo de varias coloraciones, las mitocondrias
se identifican selectivamente a travs del microscopio de
luz. La tincin de la mitocondrias se basa principalmente en
la presencia abundante de fosfolpidos membranales. La
fucsina cida empleada por Altmann y Bensley las tie de
color rojo intenso; con la hematoxilina frrica de
Heidenhain y el tetraxido de osmio se colorean de negro
intenso. La coloracin con el luxol fast blue las tie de azul
celeste ( este colorante tambin demuestra, de manera
selectiva, la mielina y el plasmalema de los eritrocitos)
Estructura electrnica de las mitocondrias. Utilizando los
microscopios electrnicos de transmisin y de barrido ha
sido posible observar imgenes sumamente detalladas de la
estructura fina de las mitocondrias. Palade, en 1953, fue el
primero en describir la ultraestructura de las mitocondrias.
Estn constituidas por los siguientes componentes:
a) Envoltura membranal. La mitocondria est limitada
por dos membranas diferentes denominadas membrana
externa y membrana interna. Esta ltima separa el contenido
mitocondrial en dos microambientes: una central o
profunda, la matriz mitocondrial y la otra perifrica, el
espacio intermembranal, situado entre la membrana interna
y la externa.
Las dos membranas difieren de forma notable tanto en su
estructura morfolgica como en la composicin de sus
componentes membranales. La membrana externa muestra
un grosor de unos 6 nm en cambio la membrana interna
suele medir de 6 a 8nm de espesor. La membrana interna
posee una mayor superficie que la externa, porque se
repliega en una serie de proyecciones llamadas crestas
mitocondriales que pueden ser laminares o tubulares. Las
crestas se disponen paralelas entre s, siguiendo el eje
mayor de la mitocondria. Las clulas que desarrollan sus
funciones con un gran gasto de ATP poseen mayor cantidad
de crestas.
La membrana externa es de superficie lisa, tiene una
composicin lipdica similar a la mayora de las membranas
celulares eucariticas, con un 30% de contenido de
colesterol. En cambio solo posee un 5% de total de las
protenas mitocondriales. Su principal protena es una
porina, que constituye canales acuosos amplios no
especficos a travs de esta membrana. Estos canales dejan
pasar molculas hidrosolubles de hasta 10 kilodaltones.
Siendo la membrana externa permeable a molculas
pequeas (especialmente protenas), el contenido del
espacio intermembranal se semeja mucho al del citoplasma.
La membrana interna difiere de manera notable en su
composicin qumica de la membrana externa. Contiene
casi el 20% de la protena mitocondrial total. Con el
microscopio electrnico y examinando superficies de
mitocondrias criofracturadas, se puede observar la presencia
de abundantes partculas protenicas insertas en la
membrana. En ella se produce la transduccin de la energa
y por el alto contenido de fosfolpidos es casi impermeable a
la mayora de iones pequeos electrones y protones. El
fosfolpido principal que alberga la membrana interna es el
bifosfatidilglicerol conocido como cardiolipina, representa
el 10% de su contenido total. La membrana externa
nicamente tiene un 3% de este fosfolpido.
En la membrana interna existen tres tipos de complejos
enzimticos:
Los componentes de la cadena respiratoria de transporte
de electrones: Complejos I, II, III y IV, el citocromo c y
la ubiquinona.
La ATP sintetasa F1 F0.
Protenas de transporte especficas, son las responsables
de regular el paso de metabolitos hacia afuera y hacia
adentro de la matriz mitocondrial.
En la superficie interna de las crestas mitocondriales, en
contacto con la matriz, Fernndez Moran, investigador
venezolano, describi en 1962 una serie de partculas
esfricas (para observarlas emple una tincin negativa con
cido fosfotungstico) de 8.0 a 9.0 nm de dimetro a las
cuales denomin partculas elementales mitocondriales.
Cada partcula se une a la cresta mitocondrial mediante un
pequeo pednculo cilndrico que mide 5nm de largo por
4nm de ancho.
Las esferas representan los lugares donde se inician los
procesos qumicos de la fosforilacin oxidativa y el
transporte de electrones. Estos palillos de tambor cortos
son complejos protenicos en los que radica la enzima
sintetasa de ATP, encargada de la gnesis de ATP a partir de
ADP y de fosfato inorgnico.
Figura Biol. Cel. 56. Fotomicrografa electrnica que muestra las
partculas de Fernndez Moran, obtenida mediante la tcnica del
sombreado negativo utilizando cido fosfotungstico.
Figura Biol. Cel. 55. a) Fotomicrografa electrnica de una mitocondria y
b) representacin esquemtica de los componentes de una mitocondria.
Las membranas mitocondriales externa e interna, en ciertas
regiones, suelen establecer contactos entre s. Estos sitios de
contacto (formados por protenas transportadoras)
intervienen como vas de transporte para protenas y
pequeas molculas que entran y salen de la matriz.
18/08/03 - 17:32 horas.
b) Espacio intermembranoso. Este espacio existe entre las
membranas externa e interna. Mide de 10 a 20 nm de ancho.
El lquido que ocupa este espacio es muy semejante, en su
composicin qumica al citosol de la clula.
c) Matriz mitocondrial. El contenido de la matriz
mitocondrial es un lquido denso, viscoso, constituido por
un 50% de protenas. La mayor parte de las protenas son
enzimas encargadas de la degradacin de los cidos grasos,
y el piruvato hasta la obtencin de la acetil CoA, con la
consiguiente oxidacin de este compuesto en el ciclo del
cido tricarboxlico (ciclo de Krebs).
En la matriz tambin se encuentran ribosomas, RNAm y
RNAt y unos grnulos densos que miden de 30 a 50 nm de
dimetro. La composicin qumica de los grnulos densos
es en base de fosfoliprotenas.
La funcin de estos grnulos no est totalmente dilucidada.
Se cree que tienen capacidad de capturar o fijar iones de
calcio y magnesio. Es notoria esta funcin en clulas seas
y cartilaginosas. Otro hecho que prueba esta funcin es
cuando en una clula lesionada, se eleva considerablemente
la concentracin de Ca2+ citoplasmtico. Los grnulos de la
matriz pueden secuestrar calcio para evitar la toxicidad que
este ion pueda producir en la clula.
En la matriz mitocondrial se localizan molculas de cido
desoxirribonucleico constituido por una doble cadena
helicoidal que se dispone de forma circular. Es un DNA no
cromosmico. Se le conoce como DNA circular (DNAc).
Similar en su presentacin al DNA de bacterias y algas
verde azules.
La existencia de RNAm, RNAt y RNAr en la matriz,
especialmente los ribosomas constituidos por subunidades
cuyos coeficientes de sedimentacin son diferentes a los
ribosomas citoslicos y se parecen ms a los que poseen las
clulas procariotas, acompaados por el DNA circular y la
presencia de enzimas para la expresin del genoma
mitocondrial, nos permite inferir que la mitocondria posee el
complejo bioqumico necesario para la sntesis de sus
propias protenas tanto membranales y enzimticas
(especialmente de la membrana interna) pero de manera
bastante restringida, pues existen evidencias que, para la
formacin y funcionalidad de las mitocondrias, se requiere
de la intervencin genmica del ncleo de la clula para
completar la sntesis de lpidos y protenas que se
incorporan, en mayor proporcin a la membrana externa que
a la interna.
Origen y reproduccin de las mitocondrias. Las
caractersticas morfolgicas y funcionales de las
mitocondrias les confieren a estos organelos al considerarlas
como componentes celulares que tienen cierta autonoma en
diversas circunstancias. La existencia de un aparato gentico
propio, de poseer los tres tipos de cido ribonucleico, y la
capacidad que tienen de originarse de otras mitocondrias, es
decir al autoreplicarse (similitud al comportamiento
bacterias y algas verde azules), apoyan la teora sustentada
por Lynn Margulis la cual afirma que durante la evolucin
de los seres vivos, la mitocondrias fueron originalmente
bacterias que invadieron o fueron fagocitadas por clulas
eucariotas, desarrollndose entre los dos organismos una
relacin de simbiosis. En la cual la futura mitocondria
recibi nutricin y proteccin de la clula husped y sta
obtuvo la capacidad de poder reducir su contenido de O 2 y
proporcionarle una forma de energa qumica permanente.
Las mitocondrias se originan por fisin o biparticin de
mitocondrias preexistentes. La
mitocondria
para
reproducirse se alarga en una forma de tubo o salchicha, en
su parte central se produce una constriccin y adopta la
forma de una clava; posteriormente se divide en dos y
origina dos mitades que despus se redondean o adquieren
una morfologa similar a la mitocondria que les dio origen.
19/08/03 16:40 horas
Funciones de las mitocondrias. Las mitocondrias
desarrollan varias funciones. La ms importantes de ellas es
la generacin de energa, especialmente la energa de trabajo
celular.
Produccin de energa. La energa se genera por la
degradacin de la glucosa y de los cidos grasos. La glucosa
es la primera sustancia que utilizan las mitocondrias para
producir energa, en caso que el suministro de glucosa se
agote, las mitocondrias emplean los cidos grasos para el
mismo fin.
La glucosa captada por las clulas es degradada
parcialmente a travs del proceso denominado gluclisis
mediante el cual despus de varios pasos la glucosa se
transforma en cido pirvico que genera pequeas
cantidades de ATP. La gluclisis se produce en el citosol y
no requiere de la presencia de oxgeno pues es un proceso
anaerobio.
El cido pirvico es captado por la mitocondria a travs de
sus membranas para llegar a la matriz, lugar donde se
transforma en acetilcoenzima A; sta a su vez se degrada en
agua (H2 O) y CO2 por la va del cido ctrico o ciclo de
Krebs.
La coenzima A tambin se produce cuando los cidos grasos
son transportados del citoplasma a la matriz mitocondrial,
mediante el proceso denominado beta-oxidacin.
19/09/03 20:55 horas
Respiracin celular. Durante el ciclo del cido ctrico o
ciclo de Krebs se liberan electrones que son transportados
por una cadena acoplada de reacciones REDOX o de
oxidorreduccin, conocida como cadena respiratoria; en
estas condiciones los electrones se unen a protones y
oxgeno, con formacin de agua. La energa generada por
este proceso oxidativo es utilizado en tres lugares para que,
a partir de ADP y fosfato inorgnico, se produzca la
regeneracin de ATP. Esta reaccin se denomina :
fosforilacin oxidativa.
mismos procesos proteolticos se producan en clulas que
carecan de lisosomas, por ejemplo las bacterias y los
eritrocitos maduros. Tenan la capacidad de destruir protena
a gran velocidad y an ms, para degradar las protenas se
necesitaba el empleo de energa a diferencia de otros
procesos degenerativos que no requeran ayuda energtica.
A fines de la dcada de los 90s, Goldberg y otros
investigadores, reportaron sus hallazgos al confirmar que
esta degradacin rpida de protenas con el uso de energa
estaba a cargo de un multicomplejo de enzimas a los cuales
se les denomin proteasomas.
Esta degradacin final de los nutrimentos aportados por los
alimentos ingeridos, que ocurre en las mitocondrias, con
consumo de oxgeno, recibe el nombre de respiracin
celular.
La gran mayora de las enzimas que intervienen en el ciclo
de Krebs se encuentran suspendidas en la matriz
mitocondrial, excepto algunas dehidrogenasas que estn
relacionadas con la membrana interna. En cambio, la cadena
transportadora de electrones se localiza en cuatro complejos
multiprotenicos localizados sobre la membrana interna.
Sntesis de esteroides. Otra de las funciones que desarrollan
las mitocondrias es aquella que, mediante enzimas
mitocondriales localizadas en la membrana interna,
catalizan la sntesis de cidos grasos y aminocidos,
especialmente la sntesis de esteroides que se inicia con la
separacin de la cadena lateral de colesterol.
El colesterol llega a las clulas, proveniente del plasma
sanguneo en la forma de una lipoprotena de baja
densidad (LDL); el compuesto que llega se encuentra como
steres de colesterol. Antes de la sntesis de esteroides se
escinden los steres del colesterol por accin de una enzima
para formar colesterol libre; de esta manera penetra el
colesterol a la mitocondria. En la membrana interna
mitocondrial existe una enzima que separa la cadena lateral
del colesterol. Este producto es transferido al retculo
endoplsmico liso en cuyo interior existen las enzimas
necesarias que completan la sntesis de esteroides.
Proteasomas. Durante los procesos de funcionales de la
clula, sta necesita digerir protenas defectuosas en su
estructura qumica o las digiere para regular una serie de
procesos importantes que involucran modificaciones o
destruccin parcial o total de algunos complejos protenicos,
que pueden ocasionar enfermedades celulares o intervenir
en la destruccin de cclicas, protenas que controlan el ciclo
celular.
La existencia de numerosas enzimas denominadas
proteasas son las responsables de tal funcin; se
consideraba que esta funcin de degradar protenas estaba a
cargo de los lisosomas, pero en los primeros aos de la
dcada de los 70s, Alfred Golberg, postul que esos
Figura Biol. Cel. 57. Representacin esquemtica de un proteasoma.
Componentes que lo integran.
Los proteasomas son grandes complejos enzimticos que
existen en todas las clulas eucariotas. Se ha demostrado
que una clula animal alberga hasta 30,000 proteasomas.
Estn integrados, como se mencion anteriormente, por
numerosas proteasas reunidas en unidades que digieren
protenas. Las proteasas adoptan la forma de cilindros
pequeos carentes de membrana que los recubra; en su
interior poseen un tnel. Cada proteasoma est constituido
por varios cilindros de proteasas.
Los proteosomas miden alrededor de 15 nm de longitud.
Los sitios proteolticos de las enzimas estn orientados hacia
la luz del tnel. Las enzimas que forman el cilindro tienen
una velocidad de sedimentacin de 20S. El cilindro est
cubierto en sus extremos por una o dos pequeas partculas
protenicas que semejan pequeos casquetes (cada uno
posee 19S). En conjunto el cilindro mas los dos casquetes
constituyen el proteasoma 26S.
Cada cilindro est constituido por cuatro anillos
helicoidales y, cada uno de ellos lo integran siete
subunidades rodeando al tnel central el cual tracto
digestivo del proteasoma. Los casquetes actan como
puertas de entrada que impediran la entrada al tnel que
protenas que, por accidente, tendran la posibilidad de
introducirse al tnel digestivo. Las partculas protenicas de
los casquetes tienen una gran selectividad para incorporar
las protenas que deben ser degradadas. Para tal fin,
utilizando energa, desenrollan la molcula protenica y la
inyectan al interior del proteasoma, lugar donde ellas sern
cortadas en partes de diverso tamao.
Este conjunto desarrolla actividad de ATPasa y reconoce
conjugados de protena a digerir y ubicuitina.
Proceso de digestin de las protenas por los proteasomas.
Las protenas que deben ser digeridas son sealadas por otra
protena la ubicuitina, denominada as porque se le ubica en
muchos organismos. La ubicuitina es una protena pequea,
formada por 76 aminocidos que se une a otras molculas
similares y constituir cadenas largas. Estas polimolculas de
ubicuitina intervienen como un cdigo seal que acelera la
entrada de la protena al proteasoma. Los procesos para la
digestin son los siguientes:
a) Existencia de tres enzimas que promueven la unin de la
protena con la ubicuitina. La primera enzima es la E1
que activa a la ubicuitina y la conecta con la enzima E2,
la enzima E3, facilita la transferencia de la ubicuitina
activada desde la enzima E2 a la protena. Este proceso
se repita hasta que la protena se encuentre unida a una
cadena larga (cola) de molculas de ubicuitina. Esta
cadena larga es reconocida por el casquete del
proteasoma y la incorpora al tnel de degradacin
protenica.
Figura Biol. Cel. 58. Esquema que representa la secuencia de eventos que
se llevan a cabo para la digestin de protenas por un proteasoma.
Citoesqueleto. Desde los inicios de la observacin de las
clulas mediante el microscopio fotnico, los cientficos
comprobaron que en el interior de las clulas existan una
serie de filamentos o pequeas trabculas que atravesaban,
en todas direcciones, o a veces de disposicin paralela, el
citoplasma celular. Inicialmente se les llam fibrillas, para
no confundirlas con las estructuras extracelulares
denominadas fibras (colgenas, elsticas y reticulares) o las
fibras musculares. De acuerdo al tipo de clula donde se
localizaban recibieron nombres especficos: neurofibrillas,
las observadas y descritas en neuronas y sus prolongaciones;
miofibrillas en las clulas musculares, tonofibrillas o
tonofilamentos en las clulas epiteliales queratinizadas, pues
se pensaba que estas ltimas le conferan el tono o la
tensin interna de las clulas para que mantuvieran su
forma.
Figura Biol. Cel. 59. Imgenes al microscopio electrnico del
citoesqueleto:
a) Citoesqueleto observado mediante el microscopio
electrnico de alto voltaje y
b) Citoesqueleto de una clula demostrado mediante el
mtodo de criofractura.
La utilizacin del microscopio electrnico y el uso de una
gran
cantidad
de
tcnicas
histoqumicas
e
inmunohistoqumicas, permiti avanzar en el conocimiento
estructural y bioqumico de estos componentes filamentosos
y las diversas funciones que cada uno de ellos desarrolla
dentro de la clula. Se comprob que la presencia de
determinados filamentos no estaba restringida a un
determinado tipo de clulas sino que tienen una distribucin
ms universal. Con el microscopio fotnico no siempre es
posible observarlos porque muchas veces estn presentes en
pequeas cantidades difciles de detectar con este tipo de
microscopio. As mismo la microscopa electrnica
demostr que las fibrillas estn constituidas por unidades
morfolgicas ms finas reconocidas como filamentos que de
manera individual, no son distinguibles con el microscopio
de luz.
Conforme se realizaron numerosos estudios con el M.E. y
las tcnicas antes descritas se dedujo que todas las clulas
eucariotas poseen esta trama fina reticulada de hilos
delgados se convino designar, al conjunto de las diversas
estructuras filamentosas contenidas en las clulas, con el
nombre genrico de citoesqueleto.
La utilizacin del microscopio electrnico de alto voltaje
( este microscopio es capaz de acelerar los electrones sobre
un potencial de un milln de voltios a diferencia del M.E
convencional que acelera los electrones varios miles de
voltios y genera un haz de electrones que atraviesan
muestras de clulas y tejidos hasta un grosor de 0.2 de
micrmetro) en la dcada de 1970, permiti observar
regiones delgadas y extendidas de clulas sin seccionar o
secciones lo suficientemente gruesas (ms de 4 o 5 m de
espesor), donde se observaba que el citoplasma contena una
red tridimensional de filamentos finos denominada trama
microtrabecular. Esta trama sirve como sustento de anclaje
y relacin entre varios de los organelos celulares e
interconecta filamentos y microtbulos. Entre la trama
microtrabecular existen los espacios intertrabeculares
llenos de contenido acuoso en el que se disuelven o
suspenden una gran cantidad de molculas pequeas que
participan en el metabolismo celular.
El citoesqueleto est constituido por cuatro tipos de
estructuras filamentosas protenicas:
Filamentos delgados. Constituidos principalmente por la
protena globular actina. Poseen un dimetro de 5 a 7 nm y
longitud variable.
Filamentos intermedios. Estn formados por varios tipos
de protenas (queratina, desmina, vimentina, lamininas,
protena cida glial). Miden en promedio 10 nm de
dimetro.
Filamentos gruesos. Inicialmente se describieron las fibras
musculares; en la actualidad su presencia se hace evidente
en todas aquellas clulas que de alguna forma desarrollan
movimientos de diversa ndole. El dimetro que poseen es
de 15 nm.
Microtbulos. Son estructuras filamentosas tubulares rectas
o a veces curvas. Estn constituidos por molculas de la
protena tubulina. Existen en casi todas las clulas
eucariotas y tambin en las plaquetas, integrantes de la
sangre, constituidas por partculas de clulas de la mdula
sea, llamadas megacariocitos. Los microtbulos miden 25
nm en su dimetro externo.
El citoesqueleto, adems, contiene numerosas protenas
accesorias que unen a los componentes y desempean
funciones fundamentales para la estructura general y
funcional del citoesqueleto. Los componentes del
citoesqueleto
intervienen de manera activa en los
movimientos intracelulares: transporte de vesculas,
partculas y organelos citoplasmticos, en el transporte de
los cromosomas durante las fases de la mitosis y en los
procesos de divisin celular.
entre s de manera helicoidal para formar un filamento
protenico (actina F) de un grosor aproximado de 7 nm y
de longitud variable (esta longitud depende del momento
funcional de la clula). Se les demostr inicialmente en las
fibras musculares; junto con los filamentos gruesos de
miosina intervienen de manera fundamental en la
contraccin
muscular.
Mediante
estudios
inmunohistoqumicos, utilizando anticuerpos contra la
protena actina, se hace evidente que existe en la gran
mayora de clulas eucariotas. Suele representar casi del 10
al 20 % del total de protenas celulares. En la mayora de
las clulas no musculares, del total de la actina presente,
solamente el 50% permanece como actina F.
La protena actina, segn su punto isoelctrico, es de tres
clases: actina alfa, propia de las clulas musculares y las
actinas beta y gamma de clulas no musculares.
Figura Biol. Cel. 61. Secuencia de la polimerizacin de las molculas
globulares de actina G, para formar cadenas de actina F y la disposicin de
las cadenas de actina F para constituir un filamento delgado.
Cuando la clula requiere de la existencia de los filamentos
de actina, se forma el filamento por acoplamiento sucesivo
de unidades globulares monomricas al extremo de un
filamento preexistente y sucede que en el otro extremo
puede generarse el desacoplamiento de los monmeros
cuando es necesario. La excepcin ocurre en las fibras
musculares, en las cuales los filamentos de actina siempre
son estables.
Figura Biol. Cel. 60. Representacin esquemtica de una clula donde se
muestran diversos integrantes del citoesqueleto.
Filamentos delgados. Llamados tambin microfilamentos,
estn constituidos por protenas globulares de actina
(actina G) que se polimerizan en dos cadenas enrolladas
Los filamentos de actina poseen polaridad con un extremo
positivo y un polo negativo. El crecimiento por
polimerizacin de molculas de actina G se produce con
mayor celeridad en el polo positivo que en el negativo. In
vitro, los filamentos de actina se ensamblan cuando los
monmeros globulares se acoplan entre s mediante la
presencia de una meromiosina, molcula integrante del
filamento grueso de miosina,
formando estructuras
filamentosas de recorrido corto en donde los extremos
positivos y los negativos se distinguen porque los negativos
adoptan la forma de puntas de flecha y los positivos de pas.
La conformacin de los filamentos de actina para constituir
el citoesqueleto o intervenir en el movimiento celular
requiere de la presencia indispensable de protenas
accesorias, denominadas protenas fijadoras de actina.
Existen dos grupos de protenas relacionadas con la actina.
Un grupo de ellas interviene en mantener el equilibrio entre
los monmeros de actina G y los filamentos de actina F
polimerizados; la protena profilina se une a los monmeros
de actina G e impide que se formen filamentos de actina F.
La brevina se une al extremo positivo de los filamentos
existentes de actina F para impedir el crecimiento del
mismo. La fragmina y la gelsolina se fijan a monmeros
aislados de actina G del filamento de actina, entre los polos
positivo y negativo de tal forma que el monmero aislado se
separa del filamento largo para escindirlo en trozos ms
pequeos. Esta ltima actividad contribuye a la degradacin
parcial del citoesqueleto y la regulacin de la longitud de los
filamentos de actina.
Otro grupo de protenas fijadoras de actina interviene en el
acoplamiento de los filamentos de actina para que
constituyan parte del citoesqueleto de la clula. Las
protenas espectrina, fodrina y filamina enlazan los
filamentos de actina por sus puntos de entrecruzamiento,
formando extensas redes que, de acuerdo a la densidad de la
trama le confiere al citosol las caractersticas de un gel
denso o laxo.
En el endoplasma (citoplasma cercano al ncleo) la trama de
actina es laxa, por lo tanto la consistencia de la regin es de
sol, en cambio por debajo del plasmalema (periferia de la
clula) la trama es ms tupida o densa y la zona posee las
caractersticas de un gel. Por ejemplo en la formacin de
seudpodos o filopodios el ensamble y desacoplamiento de
la red de actina facilita con bastante rapidez los cambios de
sol a gel y viceversa.
La villina y la fimbrina son un grupo de protenas
asociadas a filamentos de actina que se encargan de unirlos
especialmente cuando los filamentos se disponen en grupos
de orientacin paralela. De esa manera se forman haces
rgidos que discurren a travs del citoplasma confirindole
un sostn mecnico a la clula en ciertas zonas localizadas o
sirviendo de esqueleto de sostn a evaginaciones de algunas
clulas (intestinales, renales o epididimarias) conocidas
como microvellosidades.
Filamentos intermedios.
Los filamentos intermedios son estructuras protenicas
microfibrilares de longitud variable y con un grosor
aproximado de 10 nanmetros. Se les denomina intermedios
porque sus dimetros estn entre los filamentos delgados de
actina de 5 6nm y los 13 a 15 nm de los filamentos
gruesos de miosina.
22/08/03 18:00 horas
Son los elementos menos solubles del citoesqueleto, estn
considerados los componentes ms estables del
citoesqueleto, porque a un pH y fuerza inica fisiolgicos,
siempre existen en forma de filamentos o polmeros
insolubles a diferencia de los microtbulos y filamentos de
actina que son estructuras filamentosas pero que tambin
existen en forma de monmeros separados (solubles) que
dependiendo de los momentos funcionales de la clula
pueden cambiar su estructura polimrica a monomrica y
viceversa.
Los filamentos intermedios se caracterizan porque estn
constituidos por dos dobles hlices polipeptdicas
(tetrmeros) trenzadas una alrededor de la otra, para formar
un filamento protenico con giro a la izquierda.
Los filamentos intermedios existen en todas las clulas
eucariotas de invertebrados y vertebrados, algunos de ellos
se localizan especficamente en ciertas clulas. Se clasifican
en cinco tipos de acuerdo a las clases de protenas que los
integran. La disposicin especial de los polipptidos en la
constitucin de los filamentos intermedios les confiere una
gran resistencia a la tensin, por ejemplo en los filamentos
de queratina presentes en gran cantidad en las clulas
queratinizadas de la epidermis de la piel; estas clulas
ofrecen resistencia al aire, agua, bacterias y una buena parte
de sustancias qumicas.
En el cuadro anexo se enlistan los diversos tipos de
filamentos intermedios y las clulas donde existen con ms
frecuencia:
Filamentos intermedios
Clulas y tejidos
Queratina I (cida)
(tonofilamentos)
Queratina II (neutra y bsica)
(tonofilamentos)
Epiteliales
Epiteliales (uas y pelos)
Vimentina
Mesenquimatosas,
Desmina
Clulas musculares
Protena cida glial
Astrocitos, oligodendrocitos
y
clulas de
Schwann.
Neuronas (axones
y dendritas)
Neurofilamentos
Laminina
Laminas nucleares
Las funciones especficas de los filamentos intermedios no
estn an bien determinadas; en conjunto las funciones se
pueden resumir en:
Confieren sostn estructural a la clula.
Fijan al ncleo en una posicin determinada.
Constituyen un andamiaje estructural tridimensional
deformable a la clula.
Proporcionan un conjunto de conexiones adaptables
entre el plasmalema y los otros componentes del
citoesqueleto.
Forman un marco estructural para sostener la envoltura
nuclear y su consecuente reorganizacin despus de
haberse producido la mitosis.
Existen varias protenas que se adosan a los filamentos
intermedios con la finalidad de unirlos entre s. Conforme
los filamentos se unen entre ellos van integrando una red
tridimensional que favorece la estructura del citoesqueleto.
Las protenas que se relacionan a los filamentos intermedios
son:
Filagrina.
Permiten que los filamentos de queratina se asocien en
haces.
Sinamina y plectina.
Se encargan de unir filamentos de desmina y vimentina
respectivamente.
Plaquinas.
Ayudan a conservar el contacto entre los filamentos de
queratina (tonofilamentos) y los hemidesmosomas de las
clulas epiteliales para reforzar las uniones laterales y
basales de las clulas vecinas y con la membrana basal,
respectivamente. Las plaquinas tambin intervienen en
asociar filamentos de actina con los neurofilamentos de
neuronas sensoriales.
Filamentos intermedios de queratina.
Existen en todos los tipos de clulas epiteliales, las de
revestimiento o cubierta y en las glandulares (epidrmicas,
del tracto gastrointestinal, clulas hepticas y de los acinos
pancreticos). Los filamentos de queratina en las clulas
epiteliales constituyen una compleja red similar a una
canasta o cesto que rodea al ncleo y se ramifica a travs de
todo el citoplasma. Varios de los haces de filamentos
(tonofilamentos) se insertan en placas protenicas citoslicas
(plaquinas) de los desmosomas y hemidesmosomas que
intervienen en la fijacin de clulas vecinas y a la
membrana basal subepitelial. Proporcionan proteccin a las
clulas epiteliales de epitelios estratificados planos
especialmente en procesos de abrasin mecnica o de
agresin microbiana o qumica.
Neurofilamentos.
Se localizan preferentemente en las prolongaciones axnicas
de las neuronas. Se disponen en haces laxos de posicin
paralela entre s a todo lo largo del axn. Se considera que
forman parte importante de la estructura de sostn del
citoesqueleto de axones mielinizados de gran longitud.
Protena cida glial.
Sirve como soporte de las clulas gliales: astrocitos y
oligodendrocitos y clulas de Schwann.
Desmina. Estos filamentos enlazan miofibrillas de las fibras
musculares estriadas (esquelticas y cardiacas) a los discos
Z, lugar de insercin de los filamentos delgados de actina.
En las fibras musculares lisas se une a porciones densas de
actinina del citoplasma, punto de insercin de los filamentos
de actina.
Vimentina. Los filamentos intermedios de vimentina se
encuentran presentes en clulas embrionarias, y en aquellas
de origen mesenquimatoso: fibroblastos, leucocitos y
clulas endoteliales y mesoteliales. Rodean a la envoltura
nuclear localizndose en la superficie citoplasmtica de los
complejos de poro nucleares.
Laminina. Se dispone para recubrir la superficie interna de
la cubierta nuclear. Coordina el ensamble de los
componentes de la cubierta nuclear y organiza la cromatina
perinuclear.
La importancia clnica de los filamentos intermedios es que
gracias a la aplicacin de mtodos inmunohistoqumicos
especficos es posible determinar el tipo de filamentos en el
interior de clulas cancerosas que forman parte de tumores
de aparente origen desconocido.
Filamentos gruesos.
En varios libros de biologa celular y en los libros de
histologa, generalmente los filamentos gruesos no estn
considerados inicialmente dentro de la clasificacin de los
componentes fibrilares del citoesqueleto. Designan como
tales a los filamentos de miosina cuando se describen y
explican las caractersticas morfolgicas y funcionales del
sarcmero o sarcmera, la unidad contrctil de las fibras
musculares.
Los filamentos gruesos o de miosina poseen un dimetro de
15 nm o 150 amgstrons. Estn constituidos por 200 a 300
molculas de miosina (cada una de ellas tiene 150 de
largo por 2 a 3 de dimetro), organizadas en forma fibrilar
que poseen una cabeza y una cola. La cola est formada por
dos cadenas polipeptdicas de hlice alfa enrolladas entres
ellas para constituir una superhlice dextrgira que tiene
cierta rigidez. . Las cadenas en la regin de la cabeza se
separan y se extienden lateralmente en un ngulo de 80 a 90
grados enrollndose para formar una estructura globular. A
cada cabeza se les adosan dos cadenas livianas: la cadena
liviana esencial con la funcin de estabilizar la cabeza de
miosina, en tanto que la otra protena ligera que se une a la
cabeza, denominada cabeza liviana reguladora se encarga
de la regulacin de la contraccin en la fibra muscular lisa.
En cambio esta protena reguladora interviene en las fibras
musculares estriadas esqueltica y cardiaca solamente como
protena estabilizadora. Este tipo de filamento grueso
contrctil descrito se conoce como miosina II, se localiza
solamente en las clulas (fibras) musculares. En otros tipos
de clulas no musculares, la protena contrctil presente es
la molcula de miosina I que posee una sola cabeza.
Figura Biol. Cel. 62. Componentes moleculares de la miosina.
Disposicin de las molculas de miosina para formar un filamento grueso.
Utilizando enzimas proteolticas la miosina es cortada en
dos segmentos ms o menos de longitud semejante; aquella
porcin correspondiente a la cola est integrada por la
protena meromiosina ligera (MML) que carece de actividad
de ATPasa, por lo tanto no puede interactuar con la actina, la
otra protena fibrilar que forma parte importante de la
sarcmera. La otra porcin restante est integrada por la
meromiosina pesada (MMP) si posee actividad de ATPasa y
es la porcin del filamento de miosina que, durante la
contraccin del sarcmero interacciona con la actina. La
enzima papaina separa en una segunda digestin lo restante
de la cola y la cabeza.. Los filamentos gruesos estn
constituidos por varias molculas de miosina, formando un
paquete de estas protenas. Las colas se disponen paralelas
entre s y sus cabezas se proyectan a los lados a lo largo del
eje mayor. La disposicin de las cabezas se hace de forma
espaciada y escalonada a travs del eje mayor, de tal forma
que las cabezas ms distales se ponen en contacto con las
porciones iniciales de las colas (Fig. Biol. Cel. ) y as
sucesivamente, al final el filamento grueso muestra los
extremos constituidos por cabezas y porciones de las colas
entremezclados mientras que la porcin central del
filamento grueso est constituido nicamente por colas de
meromiosina ligera. Esta disposicin es la que se observa en
el complejo contrctil de las fibras musculares.
24/08/03 13:25 horas
Microtbulos.
El huso mittico, estructura fibrilar que se forma durante las
fases de la divisin celular y otros componentes del
citoesqueleto, est constituido por microtbulos. Los
microtbulos son componentes del citoesqueleto,
considerados los de mayor grosor, miden aproximadamente
25 nanmetros de dimetro, en cambio suelen mostrar
longitudes variables. En muchas clulas suelen medir unos
200 nm de longitud, en cambio en otras, como en los axones
de las neuronas pueden alcanzar hasta 25 m de longitud.
Se caracterizan porque son estructuras cilndricas, huecas
(adoptan la forma de un tubo pequeo); sus paredes miden
unos 4,5 a 5.0 nm de grosor. La luz central mide
aproximadamente 15 nm de dimetro.
Estn constituidos por dos tipos de protenas la tubulina alfa
() y la tubulina beta (). Ambas protenas poseen una
configuracin primaria semejante, lo cual indica que
provienen de un ancestro comn. Las tubulinas y son
casi idnticas en todas las clulas pero tambin guardan
caractersticas bioqumicas similares en todas las clulas
eucariotas de los seres vivos, lo que nos indica que desde
que aparecieron en clulas eucariotas no han cambiado casi
nada durante todo el proceso evolutivo de las especies. Se
puede afirmar que genticamente la tubulina es una protena
muy bien conservada.
Las molculas de tubulina alfa y beta se unen para formar
heterodmeros, los cuales se consideran las unidades
estructurales bsicas de los microtbulos.
Figura Biol. Cel. 63. Estructura molecular de los microtbulos.
Polimerizacin y ensamble de los dmeros de tubulina para
formar los protofilamentos y la integracin de 13
protofilamentos para constituir un microtbulo.
Con base en estudios bioqumicos realizados in vitro, el
empleo de tcnicas inmunohistoqumicas y el auxilio del
microscopio electrnico se ha logrado dilucidar un modelo
de organizacin de los heterodmeros de tubulina para
formar los microtbulos: Las subunidades de tubulina alfa y
beta se unen de manera espontnea para constituir los
dmeros
los
cuales
posteriormente,
si
existen
concentraciones elevadas de las subunidades se ensamblan o
se polimerizan extremo a extremo para formar cadenas que
constituyen estructuras intermedias, anillos simples o
dobles, espirales o anillos apilados.
De acuerdo a los requerimientos de la clula, estas unidades
de dmeros se abren para unirse entre s y disponerse en
forma de cadenas lineales; llamados protofilamentos. stos
se sitan unos al lado de otros hasta alcanzar un nmero de
13 protofilamentos, as constituyen lminas simples y
posteriormente unen sus lados libres hasta integrar una
estructura tubular o cilndrica simple de corta longitud.
Formado el cilindro corto se produce el crecimiento rpido
y continuo del microtbulo mediante el ensamblaje o
adicin de dmeros en un extremo del tbulo. Los
microtbulos tambin poseen polaridad de polimerizacin
de los dmeros. Tienen un extremo positivo y otro negativo.
El crecimiento del microtbulo por adicin de
es ms rpido en el polo positivo. En el polo
estabiliza la polimerizacin o ese lugar
despolimerizacin cuando as lo requiere la
microtbulo, por ejemplo.
los dmeros
negativo se
es el de
funcin del
En los centrosomas, el extremo negativo de los
microtbulos, que deben estabilizarse en su tamao, queda
embebido en la zona densa pericentriolar. En caso contrario,
en este mismo extremo negativo cuando se necesita la
reduccin del tamao de los microtbulos, se produce la
despolimerizacin de los dmeros de tubulina especialmente
durante la telofase de la mitosis en la cual los cromosomas
necesitan migrar a los polos de las futuras clulas hijas.
En las clulas animales existen lugares donde se congregan
abundantes dmeros de tubulina, por ejemplo en los
centrosomas (un par de centriolos y su matriz densa) y en
las cercanas de los cuerpos basales o cinetosomas. A partir
de estas regiones celulares se ensamblan o forman
microtbulos, por tal razn a estas regiones se les conocen
como centros de organizacin de los microtbulos
(COMT); son los encargados del ensamblaje de los dmeros
de tubulina para formar microtbulos. La polimerizacin de
las protenas y en dmeros de tubulina requiere que
stos se unan al GTP, la activacin posterior mediante la
GTP, facilita que se unan a otros dmeros o ensamblarse con
el microtbulo en formacin. Todos estos eventos de
ensamblaje se realizan en presencia de Magnesio (Mg2+).
impiden la polimerizacin porque bloquean los puntos de
ensamblaje o estando ya los microtbulos formados, los
desensamblan.
Centriolos.
En todas las clulas eucariotas animales existe en la cercana
del ncleo, una zona de citoplasma especializado llamada
centrosoma o centro celular, que contiene dos pequeos
cuerpecillos puntiformes o en forma de pequeos
bastoncitos, denominados centriolos, dispuestos cada uno
de ellos en ngulo con respecto al otro (observados con el
microscopio fotnico). Los centriolos miden 0.25 m de
dimetro por 0.25 a 2.0 m de largo. La demostracin de los
centriolos se logra cuando las clulas se colorean con
hematoxilina frrica. La posicin del centrosoma muchas
veces se observa en una pequea hendidura del ncleo,
rodeado por las cisternas de Golgi. Al par de centriolos
presente en la clula tambin se le conoce como diplosoma.
El microscopio electrnico los muestra como estructuras
cilndricas huecas de unos 400 nm de largo por 150 nm de
grosor, rodeados por un material pericentriolar amorfo y
electrondenso.
Las principales funciones de los microtbulos son:
Proporcionan rigidez y conservan la forma de la clula.
Coordinan
y
regulan
los
movimientos
intracitoplasmticos de los organelos celulares hacia
destinos determinados. Los paquetes de microtbulos
(neurotbulos) en los axones propician el transporte
axnico en ambas direcciones (anterogrado o retrogado)
de partculas, vesculas y organelos a gran velocidad (en
ciertos casos hasta 100 mm por da).
Establecen los compartimentos intracelulares no
membranosos.
Integran los componentes estructurales de cilios y
flagelos e intervienen activamente en sus movimientos.
Protenas asociadas a los microtbulos (PAM).
En la superficie externa de los microtbulos se han descrito
una serie de protenas asociadas a ellos. Se encargan de
facilitar el ensamblaje de los microtbulos, esto significa
que, en presencia de las PAM los microtbulos crecen ms
rpido y a una concentracin menor de las molculas de
tubulina; as mismo, son ms resistentes a la
despolimerizacin ya sea por accin de sustancias
desestabilizadoras como la colchicina, la vinblastina o la
vincristina, sustancias que puestas en contacto con el citosol
Figura Biol. Cel. 64. Secciones longitudinales y transversales de un
centriolo.
En secciones transversales de un centrolo se observa que la
pared est formada por nueve unidades fibrilares aplanadas,
las cuales, a su vez, estn constituidas por la unin de tres
subunidades tubulares o microtbulos que se disponen en
direccin paralela a la orientacin longitudinal del centriolo.
La triada de microtbulos recibe la designacin de
microtbulos A, B y C, conectados al centro del organelo
mediante prolongaciones radiales. Cada triada de
microtbulos adopta una ligera inclinacin angular. El
microtbulo A es el que est orientado hacia el centro y el C
ocupa una posicin perifrica. El microtbulo A es el nico
que esta constituido por 13 protofilamentos de tubulina, los
otros dos, B y C slo constan de 10 microtbulos cada uno.
21/08/03 17:00 horas
Los centriolos se duplican durante el ciclo celular, antes de
la divisin celular. En un determinado lugar del centrolo
progenitor se forma un anillo denso (Se considera que son
molculas no ensambladas de tubulina) posteriormente este
anillo se prolonga hasta formar un cilindro pequeo (el
procentriolo) en el que se empiezan a ensamblar las
molculas de tubulina para formar una pared circular con las
nueve triadas de microtbulos. Generan centriolos hijos que
se sitan perpendicularmente a los centriolos progenitores.
En esta etapa de la vida celular cada par de centriolos
(progenitor e hijo) se orientan hacia los polos de la clula y,
a partir de ellos, se ensamblan molculas de tubulina para
formar microtbulos los cuales se organizan en el huso
acromtico o mittico.
Figura Biol. Cel. 64. Fotomicrografa electrnica que muestra la relacin
que existe entre los cuerpos basales y el origen de los cilios.
Estructuras celulares semejantes a los centriolos
denominados cuerpos basales o cinetosomas son los
responsables de organizar los microtbulos que integran los
cilios y flagelos. Al igual que los centriolos, los cuerpos
basales se replican de manera independiente de la clula.
Esto significa que, a partir de los cuerpos basales hijos, se
pueden formar nuevos cilios y flagelos.
Vous aimerez peut-être aussi
- Máquinas vivientes: ¿Cómo se mueven las células?D'EverandMáquinas vivientes: ¿Cómo se mueven las células?Évaluation : 5 sur 5 étoiles5/5 (1)
- Biología Celular PDFDocument137 pagesBiología Celular PDFwestlife_zac7100% (1)
- Teoria CelularDocument21 pagesTeoria CelularValeria Alexandra De La Cruz RojasPas encore d'évaluation
- CélulaDocument30 pagesCélulamario alexanderPas encore d'évaluation
- CélulaDocument25 pagesCélulaDULCE ITZEL CAMACHO DE JESUSPas encore d'évaluation
- CelulaDocument5 pagesCelulaKaren ReyesPas encore d'évaluation
- Biologia CelularDocument2 pagesBiologia CelularWILBER OSORIO HUAIRAPas encore d'évaluation
- Biología. Conceptos Básicos /: Unidad 4. La CélulaDocument4 pagesBiología. Conceptos Básicos /: Unidad 4. La CélulaSANTIAGO TRIANA ROJAS(ESTUDIANTE)Pas encore d'évaluation
- Teoría CelularDocument3 pagesTeoría CelularYatziri MorenoPas encore d'évaluation
- Historia de La Biología CelularDocument2 pagesHistoria de La Biología Celularvanessa arpaizPas encore d'évaluation
- Celula PrimarioDocument9 pagesCelula PrimarioPALMAPas encore d'évaluation
- Célula - Wikipedia, La Enciclopedia Libre PDFDocument17 pagesCélula - Wikipedia, La Enciclopedia Libre PDFmariaPas encore d'évaluation
- Teoria Celular 2Document24 pagesTeoria Celular 2Valeria Alexandra De La Cruz RojasPas encore d'évaluation
- CélulaDocument5 pagesCélulaTomás PrietoPas encore d'évaluation
- Apuntes Teoría CelularDocument3 pagesApuntes Teoría CelularRosse Mary Olivares AedoPas encore d'évaluation
- La CelulaDocument12 pagesLa Celulafactor empresarialPas encore d'évaluation
- La CelulaDocument8 pagesLa CelulaAmy Hyleen OliveiraPas encore d'évaluation
- 05.estructura CelularDocument69 pages05.estructura Celularfrank robert rado rosalesPas encore d'évaluation
- Para Examenes Todo Sobre La CelulaDocument78 pagesPara Examenes Todo Sobre La CelulaSergio Cayra CruzPas encore d'évaluation
- Reseña Historica de La CelulaDocument10 pagesReseña Historica de La CelulaNestor J. Medina67% (3)
- Tipos CelularesDocument3 pagesTipos CelularesIngrid Vanessa Vera ArboledaPas encore d'évaluation
- Célula - Wikipedia, La Enciclopedia Libre PDFDocument133 pagesCélula - Wikipedia, La Enciclopedia Libre PDFLeonardoMgPas encore d'évaluation
- La CelulaDocument5 pagesLa CelulaMARIELIS HIDALGOPas encore d'évaluation
- La Celula Tesis FinalDocument15 pagesLa Celula Tesis FinalJhoel Antonio Quispe0% (1)
- CélulaDocument21 pagesCélulaJennifer PescioPas encore d'évaluation
- CélulaDocument24 pagesCélulafranciscoPas encore d'évaluation
- Célula TExtoDocument7 pagesCélula TExtoRosalia Reyna Torrez VillcaPas encore d'évaluation
- La CelulaDocument2 pagesLa Celulamonica alejandra guio martinezPas encore d'évaluation
- Teoria CelularDocument4 pagesTeoria CelularLorena VelazquezPas encore d'évaluation
- iNTRODUCCION A LA BIOLOGIADocument12 pagesiNTRODUCCION A LA BIOLOGIAfernanda salinasPas encore d'évaluation
- Historia de La Biología CelularDocument3 pagesHistoria de La Biología Celularnilton heiner Cerdan VasquezPas encore d'évaluation
- Evolucion de La CelulaDocument37 pagesEvolucion de La CelulaMaríaFernanda VasquezPas encore d'évaluation
- CélulaDocument14 pagesCélulaNailah22 ZhengPas encore d'évaluation
- RESEÑA HISTÓRICA DE LA BIOLOGÍA CELULAR - Castro Tapia, Víctor MartínDocument7 pagesRESEÑA HISTÓRICA DE LA BIOLOGÍA CELULAR - Castro Tapia, Víctor MartínMartinCastroTapiaPas encore d'évaluation
- La CelulaDocument6 pagesLa CelulaFabio leonel Miranda gonzalesPas encore d'évaluation
- La Célula PDFDocument3 pagesLa Célula PDFPérez Merlin Landy MelissaPas encore d'évaluation
- Teoria CelularDocument18 pagesTeoria CelularThaily UniversidadPas encore d'évaluation
- Tema 3. CélulaDocument34 pagesTema 3. CélulaDeilianny HernandezPas encore d'évaluation
- Bio Celular Gerald Karp - Facuz22Document746 pagesBio Celular Gerald Karp - Facuz22Sarah Torres100% (3)
- Biologia Celular y Molecular Gerald Karp 4ta EdicDocument746 pagesBiologia Celular y Molecular Gerald Karp 4ta EdicAdrian Aparte93% (29)
- Unidad 3 Jose YaguanaDocument61 pagesUnidad 3 Jose YaguanaNuman OrtegaPas encore d'évaluation
- CelulaDocument13 pagesCelulaalexandra leivaPas encore d'évaluation
- CélulaDocument10 pagesCélulaSofia TabordaPas encore d'évaluation
- Nuevo Documento de Microsoft WordDocument3 pagesNuevo Documento de Microsoft WordFernando Jose Castillo FernandezPas encore d'évaluation
- La CélulaDocument21 pagesLa CélulaAndrés E. Espinoza NolascoPas encore d'évaluation
- La CélulaDocument16 pagesLa CélulaRoberto AndersonPas encore d'évaluation
- CélulaDocument26 pagesCélulaTamyPas encore d'évaluation
- La Célula IntroducciónDocument4 pagesLa Célula IntroducciónYakory BallónPas encore d'évaluation
- TEMA 1 CITOLOGIA 5to - SECcundariaDocument3 pagesTEMA 1 CITOLOGIA 5to - SECcundariaGrover CopaPas encore d'évaluation
- ¿Cómo funciona una célula? Fisiología celularD'Everand¿Cómo funciona una célula? Fisiología celularÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSD'EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSPas encore d'évaluation
- Hormonas: Mensajeros químicos y comunicación celularD'EverandHormonas: Mensajeros químicos y comunicación celularÉvaluation : 2.5 sur 5 étoiles2.5/5 (2)
- Los inicios de la vida: La evolución en la Tierra precámbricaD'EverandLos inicios de la vida: La evolución en la Tierra precámbricaPas encore d'évaluation
- Genética: La continuidad de la vidaD'EverandGenética: La continuidad de la vidaÉvaluation : 3 sur 5 étoiles3/5 (1)
- Los Principios de La BiologíaDocument28 pagesLos Principios de La BiologíaKatherine MorochoPas encore d'évaluation
- Informe de Laboratorio para Microbiologia AmbientalDocument22 pagesInforme de Laboratorio para Microbiologia AmbientalCarlosDavidCelySaenzPas encore d'évaluation
- Prueba de La Célula 5 y Funciones Vitales y OrganizacionDocument4 pagesPrueba de La Célula 5 y Funciones Vitales y OrganizacionhortensiaPas encore d'évaluation
- Introducción A La Citología para Cuarto Grado de SecundariaDocument6 pagesIntroducción A La Citología para Cuarto Grado de SecundariaOsomolarPas encore d'évaluation
- Informes de BiologiaDocument20 pagesInformes de BiologiaJuan Jesus Martinez SánchezPas encore d'évaluation
- Cta4 U2Document8 pagesCta4 U2rosa luzPas encore d'évaluation
- Guia Final Bio - 10°-2022Document12 pagesGuia Final Bio - 10°-2022Carlos CardonaPas encore d'évaluation
- Nivel CelularDocument12 pagesNivel Celularyarotd sotoPas encore d'évaluation
- Ciencias de La Vida Act 14Document3 pagesCiencias de La Vida Act 14Brenda FraibPas encore d'évaluation
- Biologia 3 AduniDocument13 pagesBiologia 3 AduniHector Avendaño RamosPas encore d'évaluation
- Trabajo Biologìa HumanaDocument25 pagesTrabajo Biologìa HumanaAndresFelipeRodriguezBaqueroPas encore d'évaluation
- Partes de La CelulaDocument2 pagesPartes de La CelulaSANTIAGO CANTO ANGEL GABRIEL100% (1)
- Eaes U3-Evolución de La Vida Pb3 t6 eDocument23 pagesEaes U3-Evolución de La Vida Pb3 t6 eMiile ZambranoPas encore d'évaluation
- Evolucion de Las Bacterias - Yordy Mamani Claros - EverDocument10 pagesEvolucion de Las Bacterias - Yordy Mamani Claros - EverEver ChoquePas encore d'évaluation
- Quincena 5Document61 pagesQuincena 5Jaime Christian Jimmy Lupaca GuevaraPas encore d'évaluation
- Biologia 2 ParteDocument160 pagesBiologia 2 PartePastores Gomez DiazPas encore d'évaluation
- Adaptación CurricularDocument96 pagesAdaptación CurricularAnonymous W4Ug13aoN4100% (2)
- Biología (Celular y Molecular)Document11 pagesBiología (Celular y Molecular)David GodoyPas encore d'évaluation
- E-TEMA 1 La Microbiología y Los Microorganismos.Document28 pagesE-TEMA 1 La Microbiología y Los Microorganismos.Ester PerezPas encore d'évaluation
- Taxonomia BacterianaDocument56 pagesTaxonomia BacterianaZharick Atía DíazPas encore d'évaluation
- Biologia 6° IpDocument23 pagesBiologia 6° IpLina MedinaPas encore d'évaluation
- Actividad en CasaDocument3 pagesActividad en CasaStiven HernandezPas encore d'évaluation
- Genética MicrobianaDocument3 pagesGenética Microbianasamantha brizuelaPas encore d'évaluation
- Ensayo de BiologiaDocument53 pagesEnsayo de BiologiaOduver PerezPas encore d'évaluation
- BACTERIADocument62 pagesBACTERIALuis Molina JimenezPas encore d'évaluation
- Clasificacion de Los Seres VivosDocument51 pagesClasificacion de Los Seres Vivospacorosadesaron100% (1)
- Huellas de La Evolución Celular: La Teoría Endosimbiótica.Document32 pagesHuellas de La Evolución Celular: La Teoría Endosimbiótica.Carlos de Paz100% (2)
- ArqueobacteriasDocument7 pagesArqueobacteriasLis VeraPas encore d'évaluation
- Genética de Hongos - 1Document10 pagesGenética de Hongos - 1Gedeon CastilloPas encore d'évaluation
- BIOLOGIA-TERCERCORTE-CELULA EuCARIOTADocument14 pagesBIOLOGIA-TERCERCORTE-CELULA EuCARIOTAVictor HerreraPas encore d'évaluation