Vous aimerez peut-être aussi
- Hybrid Neural Networks: Fundamentals and Applications for Interacting Biological Neural Networks with Artificial Neuronal ModelsD'EverandHybrid Neural Networks: Fundamentals and Applications for Interacting Biological Neural Networks with Artificial Neuronal ModelsPas encore d'évaluation
- Neural Correlations, Population Coding and Computation: ReviewsDocument9 pagesNeural Correlations, Population Coding and Computation: ReviewsAlan HartogPas encore d'évaluation
- Coordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorD'EverandCoordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorJose Luis Perez VelazquezPas encore d'évaluation
- Computing With Action Potentials Roweis GOODDocument7 pagesComputing With Action Potentials Roweis GOODBill PetriePas encore d'évaluation
- Efficient Neural Codes Naturally Emerge Through Gradient Descent LearningDocument35 pagesEfficient Neural Codes Naturally Emerge Through Gradient Descent LearningPranav NyatiPas encore d'évaluation
- Neuropsicologa ConexionistaDocument38 pagesNeuropsicologa ConexionistaJenni Hernández EspinosaPas encore d'évaluation
- tmpAE4F TMPDocument10 pagestmpAE4F TMPFrontiersPas encore d'évaluation
- HD&LDocument35 pagesHD&Landerken304Pas encore d'évaluation
- Attentional Phenotypes For The Analysis of Higher Mental FunctionDocument7 pagesAttentional Phenotypes For The Analysis of Higher Mental Functionjohn fossellaPas encore d'évaluation
- Cognition AlgorithmDocument19 pagesCognition AlgorithmThiago RibeiroPas encore d'évaluation
- A Feedback Model of Visual Attention: M. W. Spratling and M. H. JohnsonDocument19 pagesA Feedback Model of Visual Attention: M. W. Spratling and M. H. JohnsonAspásiaMarianaPas encore d'évaluation
- Anatomi Dan FisiologiDocument13 pagesAnatomi Dan FisiologiPakde Putu HadiPas encore d'évaluation
- Repetition Suppression/enhancement Effects As A Result of Attractor Dynamics in Local Cortical NetworksDocument2 pagesRepetition Suppression/enhancement Effects As A Result of Attractor Dynamics in Local Cortical Networksvehaxo6855Pas encore d'évaluation
- Written Report - 10. Connectionist Approach - Johary, OmmelkhayerDocument4 pagesWritten Report - 10. Connectionist Approach - Johary, OmmelkhayerAlyanah PantaoPas encore d'évaluation
- Inferring Causality From Noninvasive Brain Stimulation in Cognitive NeuroscienceDocument29 pagesInferring Causality From Noninvasive Brain Stimulation in Cognitive NeuroscienceiNoahGH SaYHelloToOPas encore d'évaluation
- Identifying True Brain Interaction From EEG Data Using The Imaginary Part of CoherencyDocument16 pagesIdentifying True Brain Interaction From EEG Data Using The Imaginary Part of CoherencySticky NoodlePas encore d'évaluation
- PNAS 2011 Tombu 13426 31Document12 pagesPNAS 2011 Tombu 13426 31Vlad PredaPas encore d'évaluation
- Shad Len 1998 VariableDocument27 pagesShad Len 1998 VariablepastafarianboyPas encore d'évaluation
- What Is Rhythm and DotsDocument4 pagesWhat Is Rhythm and DotsAndrea Gallo de la PazPas encore d'évaluation
- Macke 2009 GeneratingDocument28 pagesMacke 2009 GeneratingpastafarianboyPas encore d'évaluation
- 1 Platonic Model of Mind As An Approximation To NeurodynamicsDocument21 pages1 Platonic Model of Mind As An Approximation To NeurodynamicsDaniel Madrid GranadosPas encore d'évaluation
- Edelman, Gerald - Neural DarwinismDocument8 pagesEdelman, Gerald - Neural DarwinismcdmemfgambPas encore d'évaluation
- #Wang Symbolic ModelDocument16 pages#Wang Symbolic ModelCombaticus001Pas encore d'évaluation
- Neocortical Dynamics at Multiple Scales: EEG Standing Wav Es, Statistical Mechanics, and Physical AnalogsDocument25 pagesNeocortical Dynamics at Multiple Scales: EEG Standing Wav Es, Statistical Mechanics, and Physical AnalogsLester IngberPas encore d'évaluation
- Common Biology of Craving Across Legal and Illegal Drugs A Quantitative Meta Analysis of Cue Ractivity Brain ResponseDocument9 pagesCommon Biology of Craving Across Legal and Illegal Drugs A Quantitative Meta Analysis of Cue Ractivity Brain ResponseJoão MaiaPas encore d'évaluation
- Relevancy of Neutrosophic Logic in Various DomainsDocument4 pagesRelevancy of Neutrosophic Logic in Various DomainsInternational Journal of Application or Innovation in Engineering & ManagementPas encore d'évaluation
- Network Structure and Dynamics of The Mental Workspace: Signi FicanceDocument6 pagesNetwork Structure and Dynamics of The Mental Workspace: Signi FicanceDavid LeónPas encore d'évaluation
- Sensory IntegrationDocument4 pagesSensory IntegrationDenise MathrePas encore d'évaluation
- Multiple Scales of Brain-Mind InteractionsDocument5 pagesMultiple Scales of Brain-Mind InteractionsLester IngberPas encore d'évaluation
- Connecting Concepts in The Brain by Mapping Cortical Representations of Semantic RelationsDocument13 pagesConnecting Concepts in The Brain by Mapping Cortical Representations of Semantic RelationsMarlon DaviesPas encore d'évaluation
- BMC Neuroscience: Assessing The Heritability of Attentional NetworksDocument7 pagesBMC Neuroscience: Assessing The Heritability of Attentional Networksjohn fossellaPas encore d'évaluation
- Neural DarwinismDocument11 pagesNeural DarwinismRodrigoPristaPas encore d'évaluation
- Conscious ConnectionDocument17 pagesConscious ConnectionProf VRShanbhagPas encore d'évaluation
- Functional and Effective Connectivity A ReviewDocument24 pagesFunctional and Effective Connectivity A ReviewIrfan Fachri MPas encore d'évaluation
- tmpBF62 TMPDocument12 pagestmpBF62 TMPFrontiersPas encore d'évaluation
- Can Emotion Modulate Attention? Evidence For Reciprocal Links in The Attentional Network TestDocument9 pagesCan Emotion Modulate Attention? Evidence For Reciprocal Links in The Attentional Network TestFrontiersPas encore d'évaluation
- Ignal Etection Heory: Instructors: J.L. Szalma and P.A. HancockDocument6 pagesIgnal Etection Heory: Instructors: J.L. Szalma and P.A. HancockLeonardoPas encore d'évaluation
- ADHD Large Scale Brain SystemsDocument10 pagesADHD Large Scale Brain SystemsBrenda FolkPas encore d'évaluation
- Hatzopoulos Hsss Education enDocument11 pagesHatzopoulos Hsss Education enJohn N HatzopoulosPas encore d'évaluation
- Keshmiri 2022Document24 pagesKeshmiri 2022Berit SingerPas encore d'évaluation
- 1 s2.0 S089662731931044X MainDocument19 pages1 s2.0 S089662731931044X MainJorge Miranda JaimePas encore d'évaluation
- On Neural Networks and Application: Submitted By: Makrand Ballal Reg No. 1025929 Mca 3 SemDocument14 pagesOn Neural Networks and Application: Submitted By: Makrand Ballal Reg No. 1025929 Mca 3 Semmakku_ballalPas encore d'évaluation
- Test YourselfDocument5 pagesTest Yourselfflay1618Pas encore d'évaluation
- Neural Manifold Analysis of Brain Circuit DynamicsDocument24 pagesNeural Manifold Analysis of Brain Circuit DynamicsKarla ValderramaPas encore d'évaluation
- Now You Hear It - Witek Vuust PDFDocument11 pagesNow You Hear It - Witek Vuust PDFAlejandro OssaPas encore d'évaluation
- A Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational PitfallsDocument23 pagesA Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational PitfallsBryan ZhengPas encore d'évaluation
- Entropy in Heart Rate Dynamics Reflects How HRV-BiDocument13 pagesEntropy in Heart Rate Dynamics Reflects How HRV-BiCKPas encore d'évaluation
- Literature Review of EegDocument6 pagesLiterature Review of Eegafmztsqbdnusia100% (1)
- s13415 017 0508 XDocument13 pagess13415 017 0508 XMarlon DaviesPas encore d'évaluation
- On The Relationships Between Generative Encodings, Regularity, and Learning Abilities When Evolving Plastic Artificial Neural NetworksDocument29 pagesOn The Relationships Between Generative Encodings, Regularity, and Learning Abilities When Evolving Plastic Artificial Neural Networksmandor-Pas encore d'évaluation
- SC SMDocument26 pagesSC SMJavi ErPas encore d'évaluation
- Towards A Statistical Mechanics of Consciousness - Guevara ErraDocument17 pagesTowards A Statistical Mechanics of Consciousness - Guevara ErraVincent MignerotPas encore d'évaluation
- Artificial Neural NetworkDocument12 pagesArtificial Neural NetworkRAZOR GAMINGPas encore d'évaluation
- Quantity Without Numbers and Numbers Without Quantity in The Parietal CortexDocument8 pagesQuantity Without Numbers and Numbers Without Quantity in The Parietal CortexRoque Lazcano VázquezPas encore d'évaluation
- 486 - Rolls++10 Choice Difficulty and Confidence in The BrainDocument13 pages486 - Rolls++10 Choice Difficulty and Confidence in The BrainAlex Ortega AlbornozPas encore d'évaluation
- Anomalous Cognition Two Protocols For DaDocument20 pagesAnomalous Cognition Two Protocols For DarammekPas encore d'évaluation
- 1 s2.0 S0959438819301102 MainDocument9 pages1 s2.0 S0959438819301102 MainAndreia SilvaPas encore d'évaluation
- Introduction To Cognitive NeuroscienceDocument7 pagesIntroduction To Cognitive Neuroscience21mpsy15Divisha Vikas ChandelPas encore d'évaluation
- Nolte G 2004 - Identifying True Brain Interaction From EEG Data Using The Imaginary Part of CoherencyDocument16 pagesNolte G 2004 - Identifying True Brain Interaction From EEG Data Using The Imaginary Part of CoherencyErik MartinezPas encore d'évaluation
- (2016) Analysis of Neuronal Spike Trains DeconstructedDocument39 pages(2016) Analysis of Neuronal Spike Trains DeconstructedDiego Soto ChavezPas encore d'évaluation
- ChurchDocument1 pageChurchLionheart GuillermoPas encore d'évaluation
- Jesus Died of A Broken HeartDocument3 pagesJesus Died of A Broken HeartLionheart GuillermoPas encore d'évaluation
- Stiffener Column Stiffener Column: Proposed Toilet and Bath Plan 7 ADocument1 pageStiffener Column Stiffener Column: Proposed Toilet and Bath Plan 7 ALionheart GuillermoPas encore d'évaluation
- 27 S-4 Typical Beam Elevation: Scale NtsDocument1 page27 S-4 Typical Beam Elevation: Scale NtsLionheart GuillermoPas encore d'évaluation
- Rear Elevation 5 A: Open Open Open OpenDocument1 pageRear Elevation 5 A: Open Open Open OpenLionheart GuillermoPas encore d'évaluation
- Proposed Assembly Hall and Toilet and Bath Plan 1 ADocument1 pageProposed Assembly Hall and Toilet and Bath Plan 1 ALionheart GuillermoPas encore d'évaluation
- AR429B PERT Final ActivityDocument32 pagesAR429B PERT Final ActivityLionheart GuillermoPas encore d'évaluation
- Tensile Structures: Pneumatic Trussed Anticlastic Stayed SuspendedDocument66 pagesTensile Structures: Pneumatic Trussed Anticlastic Stayed SuspendedLionheart GuillermoPas encore d'évaluation
- Ground Floor Plan: O T E K S T R E E TDocument1 pageGround Floor Plan: O T E K S T R E E TLionheart GuillermoPas encore d'évaluation
- Sustainable DesignDocument9 pagesSustainable DesignLionheart GuillermoPas encore d'évaluation
- Neuroanatomy QbankDocument27 pagesNeuroanatomy Qbankbbbanyi67% (3)
- Neurology of Visceral PainDocument2 pagesNeurology of Visceral PainprimzadeshaPas encore d'évaluation
- Mishkin 1983 PDFDocument4 pagesMishkin 1983 PDFamandaPas encore d'évaluation
- Laboratory ActivityDocument15 pagesLaboratory ActivityChippy RabePas encore d'évaluation
- Pain Theories PDFDocument7 pagesPain Theories PDFAndreea-Alexandra LupuPas encore d'évaluation
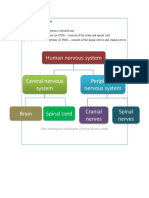
- Chart Showing The Classification of Human Nervous SystemDocument6 pagesChart Showing The Classification of Human Nervous Systemamala raviPas encore d'évaluation
- Physiological Psychology Lecture Notes: Nervous System DamageDocument3 pagesPhysiological Psychology Lecture Notes: Nervous System DamageGeneric_PersonaPas encore d'évaluation
- Response of The Brain To EnrichmentDocument66 pagesResponse of The Brain To EnrichmentCandi Kamira BalladaresPas encore d'évaluation
- Eeg EogDocument19 pagesEeg EogpranoyrajPas encore d'évaluation
- Parietal LobeDocument29 pagesParietal LobeAdrishyaPas encore d'évaluation
- Referat Patofisiologi Dan Faktor Resiko Epilepsi ParsialDocument4 pagesReferat Patofisiologi Dan Faktor Resiko Epilepsi ParsialnadiyasyadinaPas encore d'évaluation
- Ch.1 Quiz ADocument1 055 pagesCh.1 Quiz ASultan AlghamdiPas encore d'évaluation
- The Role of Early Experience in Infant DevelopmentDocument307 pagesThe Role of Early Experience in Infant DevelopmentDaru Kristiyono100% (2)
- Pain AssessmentDocument11 pagesPain AssessmentJenjen Cortey100% (1)
- Brain Storage (A Review) : Yogesh .C. Pathak, Arshad Shaikh, Akash Goswami, Himanshu Singh, Shivani Tonge, Smriti KaradeDocument3 pagesBrain Storage (A Review) : Yogesh .C. Pathak, Arshad Shaikh, Akash Goswami, Himanshu Singh, Shivani Tonge, Smriti KaradeerpublicationPas encore d'évaluation
- (Advances in Cell Aging and Gerontology 17) Mark P. Mattson (Eds.) - Sleep and Aging-Academic Press, Elsevier (2005)Document192 pages(Advances in Cell Aging and Gerontology 17) Mark P. Mattson (Eds.) - Sleep and Aging-Academic Press, Elsevier (2005)Marcos R Galvão BatistaPas encore d'évaluation
- Hebb 1949Document16 pagesHebb 1949Norel Nicolae BalutaPas encore d'évaluation
- Responses in Animals-OCR Biology A2Document17 pagesResponses in Animals-OCR Biology A2Alan TaylorPas encore d'évaluation
- Monoglutamat de SodiuDocument5 pagesMonoglutamat de SodiuadriraluPas encore d'évaluation
- Anatomi Klinik TelingaDocument51 pagesAnatomi Klinik TelingaSyahidatul ArifaPas encore d'évaluation
- Cholinergic Synaptic CommunicationDocument1 pageCholinergic Synaptic Communicationkep1313Pas encore d'évaluation
- Brain BeeDocument2 pagesBrain BeeYe Rim Won100% (1)
- JAK Neural PlasticityDocument15 pagesJAK Neural PlasticityOwais KhanPas encore d'évaluation
- Synapse, The Concept of Antidote, Curare and Exotoxins PoisoningDocument39 pagesSynapse, The Concept of Antidote, Curare and Exotoxins PoisoningCLEMENTPas encore d'évaluation
- 13 CNS PathwaysDocument17 pages13 CNS PathwaysLa Safuan AzkarPas encore d'évaluation
- Nbio 140BDocument5 pagesNbio 140Bwilliam1230Pas encore d'évaluation
- Biochemistry 1.3 NeurotransmittersDocument10 pagesBiochemistry 1.3 Neurotransmitterslovelots1234Pas encore d'évaluation
- Brodman MapDocument18 pagesBrodman MapBecky Edwards0% (1)
- Chapter 3 Subtopic 3.1 Biology (FORM 5)Document3 pagesChapter 3 Subtopic 3.1 Biology (FORM 5)AdelAutumnPas encore d'évaluation
- Stoicism The Art of Happiness: How the Stoic Philosophy Works, Living a Good Life, Finding Calm and Managing Your Emotions in a Turbulent World. New VersionD'EverandStoicism The Art of Happiness: How the Stoic Philosophy Works, Living a Good Life, Finding Calm and Managing Your Emotions in a Turbulent World. New VersionÉvaluation : 5 sur 5 étoiles5/5 (51)
- Weapons of Mass Instruction: A Schoolteacher's Journey Through the Dark World of Compulsory SchoolingD'EverandWeapons of Mass Instruction: A Schoolteacher's Journey Through the Dark World of Compulsory SchoolingÉvaluation : 4.5 sur 5 étoiles4.5/5 (149)
- Summary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisD'EverandSummary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (30)
- Follow your interests: This will make you feel better about yourself and what you can do.: inspiration and wisdom for achieving a fulfilling life.D'EverandFollow your interests: This will make you feel better about yourself and what you can do.: inspiration and wisdom for achieving a fulfilling life.Pas encore d'évaluation
- How to Improve English Speaking: How to Become a Confident and Fluent English SpeakerD'EverandHow to Improve English Speaking: How to Become a Confident and Fluent English SpeakerÉvaluation : 4.5 sur 5 étoiles4.5/5 (56)
- Summary: Trading in the Zone: Trading in the Zone: Master the Market with Confidence, Discipline, and a Winning Attitude by Mark Douglas: Key Takeaways, Summary & AnalysisD'EverandSummary: Trading in the Zone: Trading in the Zone: Master the Market with Confidence, Discipline, and a Winning Attitude by Mark Douglas: Key Takeaways, Summary & AnalysisÉvaluation : 5 sur 5 étoiles5/5 (15)
- How to Talk to Anyone: Learn the Secrets of Good Communication and the Little Tricks for Big Success in RelationshipD'EverandHow to Talk to Anyone: Learn the Secrets of Good Communication and the Little Tricks for Big Success in RelationshipÉvaluation : 4.5 sur 5 étoiles4.5/5 (1135)
- Learn Spanish While SleepingD'EverandLearn Spanish While SleepingÉvaluation : 4 sur 5 étoiles4/5 (20)
- Summary: The 5AM Club: Own Your Morning. Elevate Your Life. by Robin Sharma: Key Takeaways, Summary & AnalysisD'EverandSummary: The 5AM Club: Own Your Morning. Elevate Your Life. by Robin Sharma: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (22)
- Summary: I'm Glad My Mom Died: by Jennette McCurdy: Key Takeaways, Summary & AnalysisD'EverandSummary: I'm Glad My Mom Died: by Jennette McCurdy: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (2)
- Dumbing Us Down: The Hidden Curriculum of Compulsory SchoolingD'EverandDumbing Us Down: The Hidden Curriculum of Compulsory SchoolingÉvaluation : 4.5 sur 5 étoiles4.5/5 (498)
- The Story of the World, Vol. 1 AudiobookD'EverandThe Story of the World, Vol. 1 AudiobookÉvaluation : 4.5 sur 5 étoiles4.5/5 (3)
- The 16 Undeniable Laws of Communication: Apply Them and Make the Most of Your MessageD'EverandThe 16 Undeniable Laws of Communication: Apply Them and Make the Most of Your MessageÉvaluation : 5 sur 5 étoiles5/5 (73)
- Make It Stick by Peter C. Brown, Henry L. Roediger III, Mark A. McDaniel - Book Summary: The Science of Successful LearningD'EverandMake It Stick by Peter C. Brown, Henry L. Roediger III, Mark A. McDaniel - Book Summary: The Science of Successful LearningÉvaluation : 4.5 sur 5 étoiles4.5/5 (55)
- The Story of the World, Vol. 2 AudiobookD'EverandThe Story of the World, Vol. 2 AudiobookÉvaluation : 5 sur 5 étoiles5/5 (2)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisD'EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisÉvaluation : 5 sur 5 étoiles5/5 (3)
- Summary: Dotcom Secrets: The Underground Playbook for Growing Your Company Online with Sales Funnels by Russell Brunson: Key Takeaways, Summary & Analysis IncludedD'EverandSummary: Dotcom Secrets: The Underground Playbook for Growing Your Company Online with Sales Funnels by Russell Brunson: Key Takeaways, Summary & Analysis IncludedÉvaluation : 5 sur 5 étoiles5/5 (2)
- Cynical Theories: How Activist Scholarship Made Everything about Race, Gender, and Identity―and Why This Harms EverybodyD'EverandCynical Theories: How Activist Scholarship Made Everything about Race, Gender, and Identity―and Why This Harms EverybodyÉvaluation : 4.5 sur 5 étoiles4.5/5 (221)
- Summary: Greenlights: by Matthew McConaughey: Key Takeaways, Summary & AnalysisD'EverandSummary: Greenlights: by Matthew McConaughey: Key Takeaways, Summary & AnalysisÉvaluation : 4 sur 5 étoiles4/5 (6)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4.5 sur 5 étoiles4.5/5 (1875)
- Learn Japanese - Level 1: Introduction to Japanese, Volume 1: Volume 1: Lessons 1-25D'EverandLearn Japanese - Level 1: Introduction to Japanese, Volume 1: Volume 1: Lessons 1-25Évaluation : 5 sur 5 étoiles5/5 (17)
- Rooted in Joy: Creating a Classroom Culture of Equity, Belonging, and CareD'EverandRooted in Joy: Creating a Classroom Culture of Equity, Belonging, and CarePas encore d'évaluation
- You Are Not Special: And Other EncouragementsD'EverandYou Are Not Special: And Other EncouragementsÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- Financial Feminist: Overcome the Patriarchy's Bullsh*t to Master Your Money and Build a Life You LoveD'EverandFinancial Feminist: Overcome the Patriarchy's Bullsh*t to Master Your Money and Build a Life You LoveÉvaluation : 5 sur 5 étoiles5/5 (1)
- Little Soldiers: An American Boy, a Chinese School, and the Global Race to AchieveD'EverandLittle Soldiers: An American Boy, a Chinese School, and the Global Race to AchieveÉvaluation : 4 sur 5 étoiles4/5 (25)