Vous aimerez peut-être aussi
- 1952 Isopods in The SunDocument19 pages1952 Isopods in The SunMichael KoneznaPas encore d'évaluation
- Submitting Samples For DNA AnalysisDocument1 pageSubmitting Samples For DNA AnalysisMichael KoneznaPas encore d'évaluation
- Akeroyd PDFDocument13 pagesAkeroyd PDFThangaGiri BaskaranPas encore d'évaluation
- 2008 Tryptophan-Based Pigment in Ustilago Mol MicrobiolDocument21 pages2008 Tryptophan-Based Pigment in Ustilago Mol MicrobiolMichael KoneznaPas encore d'évaluation
- Mycotoxins in Nigerian Maize GrainsDocument9 pagesMycotoxins in Nigerian Maize Grainsmonika_ra43Pas encore d'évaluation
- MLA InTextCitationDocument13 pagesMLA InTextCitationHend KhaledPas encore d'évaluation
- 2013 Transcript Control of FUM1 Fungal Genetics and BiologyDocument8 pages2013 Transcript Control of FUM1 Fungal Genetics and BiologyMichael KoneznaPas encore d'évaluation
- Determination of GC Content of DNA With The Help of Binding Bisbenzimide 33258Document6 pagesDetermination of GC Content of DNA With The Help of Binding Bisbenzimide 33258monika_ra43Pas encore d'évaluation
- Mycotoxins in Nigerian Maize GrainsDocument9 pagesMycotoxins in Nigerian Maize Grainsmonika_ra43Pas encore d'évaluation
- Salicylic Acid Glycoside in Xylem Sap J Plant Res 2009Document9 pagesSalicylic Acid Glycoside in Xylem Sap J Plant Res 2009Michael KoneznaPas encore d'évaluation
- Mycotoxin Detoxification Deoxynivalenol 2011Document14 pagesMycotoxin Detoxification Deoxynivalenol 2011Michael KoneznaPas encore d'évaluation
- Mycotoxin Detoxification Deoxynivalenol 2011Document14 pagesMycotoxin Detoxification Deoxynivalenol 2011Michael KoneznaPas encore d'évaluation
- Endocrine Disrupters Appl Micr Biotech 2010Document13 pagesEndocrine Disrupters Appl Micr Biotech 2010Michael KoneznaPas encore d'évaluation
- Eula 50219Document5 pagesEula 50219Michael KoneznaPas encore d'évaluation
- Misuse of PCR Trends Biochem SciDocument1 pageMisuse of PCR Trends Biochem SciMichael KoneznaPas encore d'évaluation
- Misuse of PCR Trends Biochem SciDocument1 pageMisuse of PCR Trends Biochem SciMichael KoneznaPas encore d'évaluation
- Salicylic Acid Glycoside in Xylem Sap J Plant Res 2009Document9 pagesSalicylic Acid Glycoside in Xylem Sap J Plant Res 2009Michael KoneznaPas encore d'évaluation
- 2008 Int J Mol Sci - CTAB DNA ExtractionDocument16 pages2008 Int J Mol Sci - CTAB DNA ExtractionMichael KoneznaPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Practice Test 4: Explorer's Rainforest Trip and Blind AdventurerDocument6 pagesPractice Test 4: Explorer's Rainforest Trip and Blind AdventurerKhang Nguyen TienPas encore d'évaluation
- History E XDocument13 pagesHistory E XTamboli Shaikh Muaavvir AkbarPas encore d'évaluation
- Wonderland Staking and Minting Calculator (4,4)Document8 pagesWonderland Staking and Minting Calculator (4,4)Dina Maya0% (4)
- q4 Mapeh 7 WHLP Leap S.test w1 w8 MagsinoDocument39 pagesq4 Mapeh 7 WHLP Leap S.test w1 w8 MagsinoFatima11 MagsinoPas encore d'évaluation
- QDEGNSWDocument2 pagesQDEGNSWSnehin PoddarPas encore d'évaluation
- DSP Question Paper April 2012Document2 pagesDSP Question Paper April 2012Famida Begam100% (1)
- Learning CurveDocument7 pagesLearning CurveSyed FaizanPas encore d'évaluation
- Urology Semiology Guide for Renal Colic & HematuriaDocument30 pagesUrology Semiology Guide for Renal Colic & HematuriaNikita KumariPas encore d'évaluation
- List of As... SimilesDocument3 pagesList of As... SimilesFara ZahariPas encore d'évaluation
- Puregold SWOT AnalysisDocument3 pagesPuregold SWOT AnalysisRizza Mae CabigasPas encore d'évaluation
- 08.kecakapan Bahari (Indo)Document150 pages08.kecakapan Bahari (Indo)ariPas encore d'évaluation
- Culture Based Beliefs and Practices On Pregnancy and Childbirth Among Sorsoguenos Philippines PDFDocument11 pagesCulture Based Beliefs and Practices On Pregnancy and Childbirth Among Sorsoguenos Philippines PDFRazaCreciaLastrillaMenesesPas encore d'évaluation
- 00 HR Analytics 2018Document40 pages00 HR Analytics 2018Aalya ModiPas encore d'évaluation
- Certification Exam: Take A Business OnlineDocument15 pagesCertification Exam: Take A Business OnlineezerkaPas encore d'évaluation
- New Edition ThesisDocument100 pagesNew Edition Thesisniluka welagedaraPas encore d'évaluation
- BA 424 Chapter 1 NotesDocument6 pagesBA 424 Chapter 1 Notesel jiPas encore d'évaluation
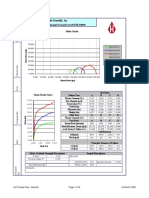
- Humboldt Scientific, Inc.: Normal Stress (Psi)Document9 pagesHumboldt Scientific, Inc.: Normal Stress (Psi)Dilson Loaiza CruzPas encore d'évaluation
- Types of Errors and Coding TechniquesDocument11 pagesTypes of Errors and Coding TechniquesTiffany KagsPas encore d'évaluation
- SM pc300350 lc6 PDFDocument788 pagesSM pc300350 lc6 PDFGanda Praja100% (2)
- Physiotherapy's Role in Rehabilitation and Health Promotion Across the LifespanDocument3 pagesPhysiotherapy's Role in Rehabilitation and Health Promotion Across the LifespanMariana OspinaPas encore d'évaluation
- Let's Meet Peter Conozcamos A PeterDocument4 pagesLet's Meet Peter Conozcamos A PeterJuan Felipe Restrepo MayaPas encore d'évaluation
- Psychotherapy Relationships That Work: Volume 1: Evidence-Based Therapist ContributionsDocument715 pagesPsychotherapy Relationships That Work: Volume 1: Evidence-Based Therapist ContributionsErick de Oliveira Tavares100% (5)
- Pathology PLE SamplexDocument5 pagesPathology PLE SamplexAileen Castillo100% (1)
- ProgDocument16 pagesProgRenon MadariPas encore d'évaluation
- God's Refining Process Removes Character DefectsDocument5 pagesGod's Refining Process Removes Character DefectsPercen7Pas encore d'évaluation
- International Human Resource Management - A Literature Review PDFDocument7 pagesInternational Human Resource Management - A Literature Review PDFTosin AdelowoPas encore d'évaluation
- Pan or The DevilDocument8 pagesPan or The DevilMarguerite and Leni Johnson100% (1)
- Certification 5Document10 pagesCertification 5juliet.clementePas encore d'évaluation
- Adeptia BPM Suite DatasheetDocument2 pagesAdeptia BPM Suite DatasheetadeptiaPas encore d'évaluation
- Tiếng Anh 6 Smart World - Unit 10 - CITIES AROUND THE WORLD - SBDocument33 pagesTiếng Anh 6 Smart World - Unit 10 - CITIES AROUND THE WORLD - SBponyoPas encore d'évaluation