Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Non-Target Effects of Clothianidin On Monarch ButterfliesDocument4 pagesNon-Target Effects of Clothianidin On Monarch ButterfliesBlue Dasher FarmPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Spatial and Numerical Relationships of Arthropod Communities Associated With Key Pests of MaizeDocument11 pagesSpatial and Numerical Relationships of Arthropod Communities Associated With Key Pests of MaizeBlue Dasher FarmPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Causes and Unintended Consequences of A Paradigm Shift in Corn Production PracticesDocument10 pagesThe Causes and Unintended Consequences of A Paradigm Shift in Corn Production PracticesBlue Dasher FarmPas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- An Indigenous Gut Bacterium, Enterococcus Faecalis (Lactobacillales: Enterococcaceae), Increases Seed Consumption by Harpalus Pensylvanicus (Coleoptera: Carabidae)Document10 pagesAn Indigenous Gut Bacterium, Enterococcus Faecalis (Lactobacillales: Enterococcaceae), Increases Seed Consumption by Harpalus Pensylvanicus (Coleoptera: Carabidae)Blue Dasher FarmPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Understanding and Enhancing Soil Biological Health: The Solution For Reversing Soil DegradationDocument40 pagesUnderstanding and Enhancing Soil Biological Health: The Solution For Reversing Soil DegradationBlue Dasher FarmPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Predator Responses To Novel Haemolymph Defences of Western Corn Rootworm (Diabrotica Virgifera) LarvaeDocument8 pagesPredator Responses To Novel Haemolymph Defences of Western Corn Rootworm (Diabrotica Virgifera) LarvaeBlue Dasher FarmPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Influence of Aphids (Myzus Persicae) and Pink Lady Beetle Larvae (Coleomegilla Maculata) On Host Plant Preference of Imported Cabbageworm (Pieris Rapae)Document8 pagesThe Influence of Aphids (Myzus Persicae) and Pink Lady Beetle Larvae (Coleomegilla Maculata) On Host Plant Preference of Imported Cabbageworm (Pieris Rapae)Blue Dasher FarmPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Trading Biodiversity For Pest Problems: ResearcharticleDocument5 pagesTrading Biodiversity For Pest Problems: ResearcharticleBlue Dasher FarmPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Mutualistic and Antagonistic Trophic Interactions in Canola: The Role of Aphids in Shaping Pest and Predator PopulationsDocument9 pagesMutualistic and Antagonistic Trophic Interactions in Canola: The Role of Aphids in Shaping Pest and Predator PopulationsBlue Dasher FarmPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- Molecular Assessment of Predation by Hoverflies (Diptera: Syrphidae) in Mediterranean Lettuce CropsDocument9 pagesMolecular Assessment of Predation by Hoverflies (Diptera: Syrphidae) in Mediterranean Lettuce CropsBlue Dasher FarmPas encore d'évaluation
- The Effects of Insecticide Dose and Herbivore Density On Tri-Trophic Effects of Thiamethoxam in A System Involving Wheat, Aphids, and LadybeetlesDocument7 pagesThe Effects of Insecticide Dose and Herbivore Density On Tri-Trophic Effects of Thiamethoxam in A System Involving Wheat, Aphids, and LadybeetlesBlue Dasher FarmPas encore d'évaluation
- Invertebrate Communities in Spring Wheat and The Identification of Cereal Aphid Predators Through Molecular Gut Content AnalysisDocument9 pagesInvertebrate Communities in Spring Wheat and The Identification of Cereal Aphid Predators Through Molecular Gut Content AnalysisBlue Dasher FarmPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Sex-Specific Interactions of Microbial Symbioses On Cricket Dietary SelectionDocument7 pagesSex-Specific Interactions of Microbial Symbioses On Cricket Dietary SelectionBlue Dasher FarmPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Spider Mite Infestations Reduce Bacillus Thuringiensis Toxin Concentration in Corn Leaves and Predators Avoid Spider Mites That Have Fed On Bacillus Thuringiensis CornDocument9 pagesSpider Mite Infestations Reduce Bacillus Thuringiensis Toxin Concentration in Corn Leaves and Predators Avoid Spider Mites That Have Fed On Bacillus Thuringiensis CornBlue Dasher FarmPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Complementarity of Statistical Treatments To Reconstruct Worldwide Routes of Invasion: The Case of The Asian Ladybird Harmonia AxyridisDocument19 pagesComplementarity of Statistical Treatments To Reconstruct Worldwide Routes of Invasion: The Case of The Asian Ladybird Harmonia AxyridisBlue Dasher FarmPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Predator Community Structure and Trophic Linkage Strength To A Focal PreyDocument9 pagesPredator Community Structure and Trophic Linkage Strength To A Focal PreyBlue Dasher FarmPas encore d'évaluation
- Protein-Marking-Based Assessment of Infield Predator DispersalDocument6 pagesProtein-Marking-Based Assessment of Infield Predator DispersalBlue Dasher FarmPas encore d'évaluation
- Molecular Determination of The Predator Community of A Cassava Whitefly in Colombia: Pest-Specific Primer Development and Field ValidationDocument7 pagesMolecular Determination of The Predator Community of A Cassava Whitefly in Colombia: Pest-Specific Primer Development and Field ValidationBlue Dasher FarmPas encore d'évaluation
- Dietary Supplementation With Pollen Enhances Survival and Collembola Boosts Fitness of A Web-Building SpiderDocument10 pagesDietary Supplementation With Pollen Enhances Survival and Collembola Boosts Fitness of A Web-Building SpiderBlue Dasher FarmPas encore d'évaluation
- Molecular Approach To Describing A Seed-Based Food Web: The Post-Dispersal Granivore Community of An Invasive PlantDocument11 pagesMolecular Approach To Describing A Seed-Based Food Web: The Post-Dispersal Granivore Community of An Invasive PlantBlue Dasher FarmPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Do Additional Sugar Sources Affect The Degree of Attendance of Dysmicoccus Brevipes by The Fire Ant Solenopsis Geminata?Document9 pagesDo Additional Sugar Sources Affect The Degree of Attendance of Dysmicoccus Brevipes by The Fire Ant Solenopsis Geminata?Blue Dasher FarmPas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- Insect Communities in Soybeans of Eastern South Dakota: The Effects of Vegetation Management and Pesticides On Soybean Aphids, Bean Leaf Beetles, and Their Natural EnemiesDocument16 pagesInsect Communities in Soybeans of Eastern South Dakota: The Effects of Vegetation Management and Pesticides On Soybean Aphids, Bean Leaf Beetles, and Their Natural EnemiesBlue Dasher FarmPas encore d'évaluation
- Effects of Neonicitinoid Seed Treatments On Soybean Aphid and Its Natural EnemiesDocument8 pagesEffects of Neonicitinoid Seed Treatments On Soybean Aphid and Its Natural EnemiesBlue Dasher FarmPas encore d'évaluation
- RNAi-Based Insecticidal Crops: Potential Effects On Nontarget SpeciesDocument9 pagesRNAi-Based Insecticidal Crops: Potential Effects On Nontarget SpeciesBlue Dasher FarmPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Why Eat Extrafloral Nectar? Understanding Food Selection by Coleomegilla Maculata (Coleoptera: Coccinellidae)Document11 pagesWhy Eat Extrafloral Nectar? Understanding Food Selection by Coleomegilla Maculata (Coleoptera: Coccinellidae)Blue Dasher FarmPas encore d'évaluation
- Insecticide Use and Crop Selection in Regions With High GM Adoption RatesDocument11 pagesInsecticide Use and Crop Selection in Regions With High GM Adoption RatesBlue Dasher FarmPas encore d'évaluation
- Functional Responses To Food Diversity: The Effect of Seed Availability On The Feeding Behavior of Facultative GranivoresDocument9 pagesFunctional Responses To Food Diversity: The Effect of Seed Availability On The Feeding Behavior of Facultative GranivoresBlue Dasher FarmPas encore d'évaluation
- Effect of Prior Diet On Consumption and Digestion of Prey and Non-Prey Food by Adults of The Generalist Predator Coleomegilla MaculataDocument7 pagesEffect of Prior Diet On Consumption and Digestion of Prey and Non-Prey Food by Adults of The Generalist Predator Coleomegilla MaculataBlue Dasher FarmPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Consumer Behaviour: Group ProjectDocument5 pagesConsumer Behaviour: Group ProjectAanchal MahajanPas encore d'évaluation
- Genetics ProblemsDocument50 pagesGenetics ProblemsTasneem SweedanPas encore d'évaluation
- English Financial Assistance ApplicationDocument4 pagesEnglish Financial Assistance ApplicationAlyssa JenningsPas encore d'évaluation
- FileDocument284 pagesFileJesse GarciaPas encore d'évaluation
- Hinduism Today April May June 2015Document43 pagesHinduism Today April May June 2015jpmahadevPas encore d'évaluation
- Ic Audio Mantao TEA2261Document34 pagesIc Audio Mantao TEA2261EarnestPas encore d'évaluation
- Lease Practice QuestionsDocument4 pagesLease Practice QuestionsAbdul SamiPas encore d'évaluation
- Case Report on Right Knee FuruncleDocument47 pagesCase Report on Right Knee Furuncle馮宥忻Pas encore d'évaluation
- 2016.05.16 - Org ChartDocument2 pages2016.05.16 - Org ChartMelissaPas encore d'évaluation
- Slaked Lime MSDS Safety SummaryDocument7 pagesSlaked Lime MSDS Safety SummaryFurqan SiddiquiPas encore d'évaluation
- Stress and FilipinosDocument28 pagesStress and FilipinosDaniel John Arboleda100% (2)
- Ignition System Spark Test DiagnosisDocument24 pagesIgnition System Spark Test DiagnosisMohamed l'Amine75% (4)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Reading and Listening 2Document4 pagesReading and Listening 2Hải Anh TạPas encore d'évaluation
- QRF HD785-7Document2 pagesQRF HD785-7Ralf MaurerPas encore d'évaluation
- Funds Flow Statement ExplainedDocument76 pagesFunds Flow Statement Explainedthella deva prasad0% (1)
- Vturn-NP16 NP20Document12 pagesVturn-NP16 NP20José Adalberto Caraballo Lorenzo0% (1)
- Goals Editable PDFDocument140 pagesGoals Editable PDFManuel Ascanio67% (3)
- Grab Go Porter S 5 ForcesDocument2 pagesGrab Go Porter S 5 ForcesUtkarsh SharmaPas encore d'évaluation
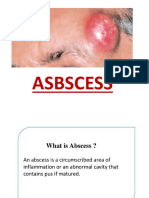
- ABSCESSDocument35 pagesABSCESSlax prajapatiPas encore d'évaluation
- Social Studies SbaDocument12 pagesSocial Studies SbaSupreme KingPas encore d'évaluation
- COVID 19 Impacts On The Construction IndustryDocument46 pagesCOVID 19 Impacts On The Construction IndustryAlemayehu DargePas encore d'évaluation
- Siemens MV Gas Insulated Switch GearDocument14 pagesSiemens MV Gas Insulated Switch GearSajesh Thykoodan T VPas encore d'évaluation
- Life Overseas 7 ThesisDocument20 pagesLife Overseas 7 ThesisRene Jr MalangPas encore d'évaluation
- Valvula de Leve MasterDocument20 pagesValvula de Leve Masterguillermo trejosPas encore d'évaluation
- Alternate Dialysis Platforms:: Sorbents SorbentsDocument17 pagesAlternate Dialysis Platforms:: Sorbents SorbentsJoe Single100% (2)
- Urban Drainage Modelling Guide IUD - 1Document196 pagesUrban Drainage Modelling Guide IUD - 1Helmer Edgardo Monroy GonzálezPas encore d'évaluation
- Book 1Document94 pagesBook 1JOHNPas encore d'évaluation
- Dip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Document1 pageDip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Neo Latoya MadunaPas encore d'évaluation
- Immune System Quiz ResultsDocument6 pagesImmune System Quiz ResultsShafeeq ZamanPas encore d'évaluation
- Chemical and Physical Properties of Refined Petroleum ProductsDocument36 pagesChemical and Physical Properties of Refined Petroleum Productskanakarao1Pas encore d'évaluation
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessD'Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessÉvaluation : 4 sur 5 étoiles4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceD'EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceÉvaluation : 4.5 sur 5 étoiles4.5/5 (515)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeD'EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifePas encore d'évaluation
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindD'EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindÉvaluation : 4.5 sur 5 étoiles4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondD'EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondÉvaluation : 4 sur 5 étoiles4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)