Vous aimerez peut-être aussi
- Evaluacion NutricionalDocument3 pagesEvaluacion NutricionalAnthonyKleinNuñezAlarconPas encore d'évaluation
- Cancer Laboral JCyL PDFDocument144 pagesCancer Laboral JCyL PDFAnthonyKleinNuñezAlarconPas encore d'évaluation
- TuberculosisDocument6 pagesTuberculosisAnthonyKleinNuñezAlarcon100% (1)
- Evolucion03 100418065842 Phpapp02Document29 pagesEvolucion03 100418065842 Phpapp02Maka Gomez Ibarra100% (2)
- Caso ClinicoDocument1 pageCaso ClinicoEdisonAlonzoPazPas encore d'évaluation
- Transaminacion 2017Document19 pagesTransaminacion 2017AnthonyKleinNuñezAlarcon0% (1)
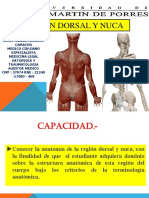
- Clase Nuca y Region DorsalDocument53 pagesClase Nuca y Region DorsalAnthonyKleinNuñezAlarconPas encore d'évaluation
- Parasito 3.1Document17 pagesParasito 3.1AnthonyKleinNuñezAlarconPas encore d'évaluation
- Biosintesis AminoacidosDocument63 pagesBiosintesis AminoacidosAnthonyKleinNuñezAlarconPas encore d'évaluation
- HepatitisDocument41 pagesHepatitisAnthonyKleinNuñezAlarconPas encore d'évaluation
- Clasificacion de Riesgo de Dispositivos MedicosDocument3 pagesClasificacion de Riesgo de Dispositivos Medicosmaylee smith sandoval100% (1)
- Rinitis Crnica AlrgicaDocument9 pagesRinitis Crnica AlrgicananPas encore d'évaluation
- Bases Farmacologicas Del Tratamiento de Las Queratitis Bacterianas PDFDocument5 pagesBases Farmacologicas Del Tratamiento de Las Queratitis Bacterianas PDFJose Antonio Fuentes VegaPas encore d'évaluation
- Malformaciones Congénitas Del Sistema NerviosoDocument5 pagesMalformaciones Congénitas Del Sistema NerviosoMartin SanchezPas encore d'évaluation
- Edema Pulmonar No CardiogénicoDocument35 pagesEdema Pulmonar No CardiogénicoIrlanda Maldonado100% (1)
- Exposición Control de La Expresión Genética y RibointerruptoresDocument2 pagesExposición Control de La Expresión Genética y RibointerruptoresAnnerys EscañoPas encore d'évaluation
- Las Etapas Del EmbarazoDocument3 pagesLas Etapas Del EmbarazoDoble ClickPas encore d'évaluation
- Diabetes Negocio MortalDocument12 pagesDiabetes Negocio MortalCésar Paz100% (1)
- 16 Rippe El CicloEstral FinalDocument6 pages16 Rippe El CicloEstral FinalJorge CampilloPas encore d'évaluation
- Bio Magnetism oDocument29 pagesBio Magnetism omisabelfuente3164Pas encore d'évaluation
- Zombiología v1.0Document28 pagesZombiología v1.0Daniel Magariño BarretoPas encore d'évaluation
- Las Plantas Magicas y La Conciencia VisionariaDocument9 pagesLas Plantas Magicas y La Conciencia Visionariadiego_griffon100% (3)
- CP Micro2Document51 pagesCP Micro2Gerardo FloresPas encore d'évaluation
- Rayos UVDocument18 pagesRayos UVIngridIrisMelgarMoraPas encore d'évaluation
- La Criminalística IvDocument37 pagesLa Criminalística IvMorrissey100% (5)
- Embriologia General.Document9 pagesEmbriologia General.Candela Carrion BustosPas encore d'évaluation
- Como Descalcificar y Desintoxicar La Glándula PinealDocument6 pagesComo Descalcificar y Desintoxicar La Glándula PinealcesarPas encore d'évaluation
- Manual Toma MuestraDocument48 pagesManual Toma MuestraJalea HumanaPas encore d'évaluation
- Cancer GastricoDocument21 pagesCancer GastricoYezzi Arguello RamosPas encore d'évaluation
- Experimento de Mucosa LabialDocument2 pagesExperimento de Mucosa LabialCRISTHIAN EDUARDO IMAN IMANPas encore d'évaluation
- Seminario MamasDocument10 pagesSeminario MamasFabiana QuinteroPas encore d'évaluation
- Cavalli-Sforza Quiénes Somos Cap. 5 en Cuanto Nos Diferenciamos Historia Genética de La HumanidadDocument2 pagesCavalli-Sforza Quiénes Somos Cap. 5 en Cuanto Nos Diferenciamos Historia Genética de La HumanidadnatyPas encore d'évaluation
- Drenaje LinfaticoDocument15 pagesDrenaje LinfaticoLeonel Dante PoloPas encore d'évaluation
- MiedoDocument4 pagesMiedoMichel OlivaresPas encore d'évaluation
- Manual de Diagnóstico PsiquiátricoDocument24 pagesManual de Diagnóstico PsiquiátricoArturo ChávezPas encore d'évaluation
- Cabalgando en Un Silbido. Omar ViñoleDocument150 pagesCabalgando en Un Silbido. Omar ViñoleGeo NautaPas encore d'évaluation
- Trabajo SoledadDocument1 pageTrabajo SoledadIvan FajardoPas encore d'évaluation
- Unidad de La VidaDocument19 pagesUnidad de La VidaMaría beronica Beltre tejadaPas encore d'évaluation