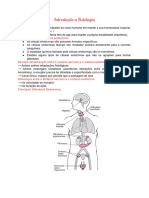
Vous aimerez peut-être aussi
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoD'EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoPas encore d'évaluation
- Rejuvenescer Com O Plasma Sanguíneo Dos JovensD'EverandRejuvenescer Com O Plasma Sanguíneo Dos JovensPas encore d'évaluation
- Questoes BioquimicaDocument4 pagesQuestoes BioquimicalarisabdinPas encore d'évaluation
- Tema 4 - Fisiologia Do Sistema HormonalDocument35 pagesTema 4 - Fisiologia Do Sistema HormonalFlávia Couto LealPas encore d'évaluation
- Hormonios BrigoniDocument11 pagesHormonios BrigoniPatricia Ferais FeraisPas encore d'évaluation
- 02 - Classificação Dos HormôniosDocument3 pages02 - Classificação Dos HormôniosLuis OliveiraPas encore d'évaluation
- Coordenação Das Funções Corporais Por Mensageiros QuímicosDocument7 pagesCoordenação Das Funções Corporais Por Mensageiros QuímicosFernando Baptista Felipe Leão LeãoPas encore d'évaluation
- Comunicação CelularDocument5 pagesComunicação CelularAndré Mahmoud100% (1)
- Estresse, pressão arterial e glicemiaDocument8 pagesEstresse, pressão arterial e glicemiaImagine WriterPas encore d'évaluation
- Fisiologia EndócrinaDocument31 pagesFisiologia EndócrinaJuliana CampbellPas encore d'évaluation
- Estudo Dirigido MachosDocument4 pagesEstudo Dirigido MachosPEDRO HENRIQUE JORENTIPas encore d'évaluation
- Cap 74 - Introdução A EndocrinologiaDocument7 pagesCap 74 - Introdução A Endocrinologiaisabella dacrocePas encore d'évaluation
- 04 ProteinasDocument5 pages04 ProteinasSamuelSamuelPas encore d'évaluation
- Estudo Dirigido de Fisiologia Endócrina Luciana Gentile GabaritoDocument7 pagesEstudo Dirigido de Fisiologia Endócrina Luciana Gentile GabaritoWellington SilvaPas encore d'évaluation
- Comunicação Celular por Sinais QuímicosDocument2 pagesComunicação Celular por Sinais QuímicosMaria Theresa MoreiraPas encore d'évaluation
- Hormone Biosynthesis, Metabolism, and Mechanism ofDocument42 pagesHormone Biosynthesis, Metabolism, and Mechanism ofLeonardo BezerraPas encore d'évaluation
- Defeitos no transportador de glicose e diabetesDocument7 pagesDefeitos no transportador de glicose e diabetesGabriela LopesPas encore d'évaluation
- 995 - Aula de Endocrinologia Da ReproduçãoDocument46 pages995 - Aula de Endocrinologia Da ReproduçãoVictor AmorimPas encore d'évaluation
- Lab Morfo UC10 - Aula 01 - Mecanismos de Ação HormonalDocument31 pagesLab Morfo UC10 - Aula 01 - Mecanismos de Ação HormonalIsa SilvaPas encore d'évaluation
- Bioquímicas IntroduçãoDocument23 pagesBioquímicas Introdução23003186Pas encore d'évaluation
- ENDOCRINOLOGIADocument58 pagesENDOCRINOLOGIADanielle MariaPas encore d'évaluation
- Fisiologia Ciclo 3Document6 pagesFisiologia Ciclo 3Fabio MorenoPas encore d'évaluation
- Caio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasDocument4 pagesCaio Maximus - Primeira Avaliação Do Curso de Biofísica Celular e Dos SistemasCaio MaximusPas encore d'évaluation
- Guião CBL 1Document53 pagesGuião CBL 1Diana BatistaPas encore d'évaluation
- GABARITODocument6 pagesGABARITOMarcio CastroPas encore d'évaluation
- Fisiologia - ProvaDocument32 pagesFisiologia - ProvaEMILY MIKELY SILVA DE MELOPas encore d'évaluation
- Questionário - Endocrinologia FURG - GabaritoDocument4 pagesQuestionário - Endocrinologia FURG - GabaritoTaironePas encore d'évaluation
- INTRODUÇÃO A FISIOLOGIA (Biologia Celular)Document19 pagesINTRODUÇÃO A FISIOLOGIA (Biologia Celular)geojac2112Pas encore d'évaluation
- Síntese e catabolismo de proteínasDocument13 pagesSíntese e catabolismo de proteínasLuis Mateus Camelo França MartinsPas encore d'évaluation
- Comunicação e Sinalização CelularDocument7 pagesComunicação e Sinalização CelularAna Resende100% (1)
- Controle da expressão génica em diferentes níveisDocument15 pagesControle da expressão génica em diferentes níveisEsm KátyaPas encore d'évaluation
- 5.1. Sistema EndócrinoDocument46 pages5.1. Sistema EndócrinoNatália Oiring100% (1)
- Secreçao HormonalDocument21 pagesSecreçao HormonalFernando Baptista Felipe Leão LeãoPas encore d'évaluation
- Resumo - Calcitonina - Camila Borges ATM 2022Document3 pagesResumo - Calcitonina - Camila Borges ATM 2022TaironePas encore d'évaluation
- Vias sinalização expressão gênicaDocument8 pagesVias sinalização expressão gênicaJoao ManoelPas encore d'évaluation
- Fisiologia do Sistema Endócrino: Funções, Hormônios e ControleDocument7 pagesFisiologia do Sistema Endócrino: Funções, Hormônios e ControlePhelipe PessanhaPas encore d'évaluation
- Relatório Sistema EndócrinoDocument32 pagesRelatório Sistema EndócrinoRogerMelo0% (1)
- Integração MetabólicaDocument35 pagesIntegração MetabólicaRicardo BorgesPas encore d'évaluation
- Castração e Terapêutica Hormonal em RatosDocument20 pagesCastração e Terapêutica Hormonal em RatosPersio M. OliveiraPas encore d'évaluation
- Jornal Da Cidade Post Feed para Instag32443E Preto23Document20 pagesJornal Da Cidade Post Feed para Instag32443E Preto23Lucas HenriquePas encore d'évaluation
- Introdução ao Sistema EndócrinoDocument11 pagesIntrodução ao Sistema Endócrinomatheusfed2Pas encore d'évaluation
- Resumão Fisiologia Dos HormôniosDocument13 pagesResumão Fisiologia Dos HormôniosEstefany CamilaPas encore d'évaluation
- Problema 8Document12 pagesProblema 8Moises Popp de OliveiraPas encore d'évaluation
- A Função Hormonal No Metabolismo CelularDocument11 pagesA Função Hormonal No Metabolismo CelularRondinele MouttaPas encore d'évaluation
- Apostila de Fisiologia Aplicada PDFDocument10 pagesApostila de Fisiologia Aplicada PDF12ab34cd8969Pas encore d'évaluation
- Introdução A FisiologiaDocument4 pagesIntrodução A Fisiologializ de andradePas encore d'évaluation
- 1 AP Fisiologia 2020.2Document8 pages1 AP Fisiologia 2020.2Mari SousaPas encore d'évaluation
- Pricipais NeurotransmissoresDocument9 pagesPricipais NeurotransmissoresTarssila Rafaela Santos88% (8)
- Sistemas neuroendócrino e digestivoDocument10 pagesSistemas neuroendócrino e digestivoFernando GuerraPas encore d'évaluation
- Neurotransmissores e ReceptoresDocument9 pagesNeurotransmissores e ReceptoresJennifer Fontenelle100% (1)
- Unidade II - Tema 3 - ProteínasDocument15 pagesUnidade II - Tema 3 - ProteínasINSTITUTO DE ENSINO INESPECPas encore d'évaluation
- Os principais hormônios e suas funçõesDocument38 pagesOs principais hormônios e suas funçõesKalina SousaPas encore d'évaluation
- Aula 10 - Drogas Colinérgicas Ou ParassimpáticomiméticasDocument6 pagesAula 10 - Drogas Colinérgicas Ou ParassimpáticomiméticasGiselli Rodrigues CordeiroPas encore d'évaluation
- Fisiologia Celular e Sistema NervosoDocument18 pagesFisiologia Celular e Sistema NervosoFernan DaviPas encore d'évaluation
- Exercício de fixação sobre anatomia, fisiologia e desenvolvimento embrionárioDocument9 pagesExercício de fixação sobre anatomia, fisiologia e desenvolvimento embrionárioMiguel LisboaPas encore d'évaluation
- Farmacologia - AnticoncepcionaisDocument20 pagesFarmacologia - Anticoncepcionaiskahionz100% (2)
- Transdução de SinaisDocument6 pagesTransdução de SinaisthuanesgPas encore d'évaluation
- Sinal CelularDocument22 pagesSinal CelularBruno CordeiroPas encore d'évaluation
- Formação Da TireoideDocument3 pagesFormação Da TireoideLaise Karolina Torres da SilvaPas encore d'évaluation
- Questões FisiologiaDocument6 pagesQuestões FisiologiaPaulo Eduardo Souza de MelosPas encore d'évaluation
- Cálculo IDHDocument4 pagesCálculo IDHLucas MarquesPas encore d'évaluation
- Ansiolíticos - Tratamento e mecanismos de açãoDocument8 pagesAnsiolíticos - Tratamento e mecanismos de açãoLucas Marques100% (1)
- Lista de Exercícios 1 Tec 1 2017.1Document3 pagesLista de Exercícios 1 Tec 1 2017.1Lucas MarquesPas encore d'évaluation
- (08.29 e 08.30) SNA ColinergicoDocument65 pages(08.29 e 08.30) SNA ColinergicoLucas MarquesPas encore d'évaluation
- case2: DOCUMENTODocument4 pagescase2: DOCUMENTOLucas MarquesPas encore d'évaluation
- Técnico - QuimioterápicosDocument30 pagesTécnico - QuimioterápicosLucas MarquesPas encore d'évaluation
- Manual Endocrinologia Julho13 FINALDocument116 pagesManual Endocrinologia Julho13 FINALCelicio DomingosPas encore d'évaluation
- Trabalho Sistema EndócrinoDocument2 pagesTrabalho Sistema EndócrinoBruna VieiraPas encore d'évaluation
- Sistema endócrino: hormônios da tireoide e distúrbiosDocument50 pagesSistema endócrino: hormônios da tireoide e distúrbiosandre almeidaPas encore d'évaluation
- Np3 1mutações No Câncer - ResumoDocument10 pagesNp3 1mutações No Câncer - ResumoCaroline MesquitaPas encore d'évaluation
- Ultrassonografia abdominal em pequenos animaisDocument81 pagesUltrassonografia abdominal em pequenos animaisRose AlvesPas encore d'évaluation
- Pop - IntradermoDocument4 pagesPop - IntradermoCintia DantasPas encore d'évaluation
- Lise Tumoral MOC BrasilDocument4 pagesLise Tumoral MOC BrasilLuciano LaranjeiraPas encore d'évaluation
- Biomoléculas e metabolismo celularDocument4 pagesBiomoléculas e metabolismo celularMariana RizzatoPas encore d'évaluation
- Função renal e os principais examesDocument36 pagesFunção renal e os principais examesLuíze FoizerPas encore d'évaluation
- Maria Tereza Beltrao Silva de Medeiros 6650691631 13/06/2023Document9 pagesMaria Tereza Beltrao Silva de Medeiros 6650691631 13/06/2023LinoPas encore d'évaluation
- 2o-DIA-PROVA - 2022-2Document12 pages2o-DIA-PROVA - 2022-2Rebeca LucioPas encore d'évaluation
- APOSTILA 3 - Bioquimica - Biomoleculas Conc. e Met. - BIO. 2018-53a400 buBT7kKDocument105 pagesAPOSTILA 3 - Bioquimica - Biomoleculas Conc. e Met. - BIO. 2018-53a400 buBT7kKLaisa MendesPas encore d'évaluation
- Resultados de exames de Glauco RobertoDocument6 pagesResultados de exames de Glauco RobertoGLÁUCIA FERNANDA CERON CERONPas encore d'évaluation
- Dicionário semiológico da UNIRIODocument13 pagesDicionário semiológico da UNIRIOTriz BeaPas encore d'évaluation
- Metabolismo de lipídeos: beta-oxidação e cetogêneseDocument7 pagesMetabolismo de lipídeos: beta-oxidação e cetogênesejose ribamar silva dos santosPas encore d'évaluation
- Bioquímica Clínica Resumo 2Document6 pagesBioquímica Clínica Resumo 2Vitor PontesPas encore d'évaluation
- Códigos sagrados para saúde física, emocional, mental e espiritualDocument22 pagesCódigos sagrados para saúde física, emocional, mental e espiritualSonia De Almeida50% (2)
- Trabalho Farmacologia Aplicada Mapa Conceitual Sobre Fármacos Utilizados No Tratamento Da AIDSDocument1 pageTrabalho Farmacologia Aplicada Mapa Conceitual Sobre Fármacos Utilizados No Tratamento Da AIDSRayssa Maria Tapajos CoqueiroPas encore d'évaluation
- Morte celular: necrose e apoptoseDocument49 pagesMorte celular: necrose e apoptoseprietoml50% (2)
- 5.1. Sistema EndócrinoDocument46 pages5.1. Sistema EndócrinoNatália Oiring100% (1)
- Resultados de exames laboratoriaisDocument3 pagesResultados de exames laboratoriaisRô SilvaPas encore d'évaluation
- Atividade física para idosos: benefícios e orientaçõesDocument22 pagesAtividade física para idosos: benefícios e orientaçõesTamires BragaPas encore d'évaluation
- CM - EndócrinoDocument44 pagesCM - EndócrinoQuênia AlmeidaPas encore d'évaluation
- Aula 10 - Bioquímica de Alimentos de Origem AnimalDocument65 pagesAula 10 - Bioquímica de Alimentos de Origem Animalgabriel lucas marques de souzaPas encore d'évaluation
- Morte celular: tipos de necroseDocument51 pagesMorte celular: tipos de necroseRaylane SilvaasPas encore d'évaluation
- Sistema endócrino e suas glândulasDocument44 pagesSistema endócrino e suas glândulasbarb100% (1)
- Exames médicos prescritos Dra. Ariane CardosoDocument3 pagesExames médicos prescritos Dra. Ariane CardosoPriscila ThaísPas encore d'évaluation
- Sais MineraisDocument6 pagesSais MineraisCelicio Domingos DomingosPas encore d'évaluation
- Sistemas endócrino e nervoso: estruturas, funções e controleDocument46 pagesSistemas endócrino e nervoso: estruturas, funções e controleDavi Barcelos SuttoffPas encore d'évaluation
- Respostas Fisiológicas e Da Performance em Exercícios de Corrida em Intensidades Máxima e SupramáximaDocument55 pagesRespostas Fisiológicas e Da Performance em Exercícios de Corrida em Intensidades Máxima e SupramáximaDiego UrsiniPas encore d'évaluation