Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Ma Tri Lineal ComponentsDocument6 pagesMa Tri Lineal Componentscverret311Pas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Mulitiple Maternal Origins - Goes With Ma Tri Lineal ComponentsDocument8 pagesMulitiple Maternal Origins - Goes With Ma Tri Lineal Componentscverret311Pas encore d'évaluation
- Growth PerformanceDocument9 pagesGrowth Performancecverret311Pas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Effect of Dietary EnergyDocument5 pagesEffect of Dietary Energycverret311Pas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Comparison of Different Ideal AA RatiosDocument5 pagesComparison of Different Ideal AA Ratioscverret311Pas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Chemical CompositionsDocument8 pagesChemical Compositionscverret311Pas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Arbor Acres Broiler ManualDocument64 pagesArbor Acres Broiler Manualcverret311100% (5)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Steinecker Boreas: Wort Stripping of The New GenerationDocument16 pagesSteinecker Boreas: Wort Stripping of The New GenerationAlejandro Javier Delgado AraujoPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Checklist Code ReviewDocument2 pagesChecklist Code ReviewTrang Đỗ Thu100% (1)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Balkhu Squatter SettlementDocument10 pagesBalkhu Squatter SettlementShramina ShresthaPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Research Article: Dynamic Modelling of A Solar Water Pumping System With Energy StorageDocument13 pagesResearch Article: Dynamic Modelling of A Solar Water Pumping System With Energy Storagehabtamu melsewPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- SoC Showdown: Snapdragon 810 Vs Exynos 7420 Vs MediaTek Helio X10 Vs Kirin 935 - AndroidAuthority - PDDocument19 pagesSoC Showdown: Snapdragon 810 Vs Exynos 7420 Vs MediaTek Helio X10 Vs Kirin 935 - AndroidAuthority - PDArpit SharmaPas encore d'évaluation
- Defence QuestionnaireDocument2 pagesDefence QuestionnaireSumitt SinghPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- FixDocument4 pagesFixReza FahmiPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- BTL Marketing CompanyDocument30 pagesBTL Marketing Companypradip_kumarPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- HOWO SERVICE AND MAINTENANCE SCHEDULE SinotruckDocument3 pagesHOWO SERVICE AND MAINTENANCE SCHEDULE SinotruckRPaivaPas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- VSL Synchron Pianos Changelog en 1.1.1386Document4 pagesVSL Synchron Pianos Changelog en 1.1.1386RdWingPas encore d'évaluation
- Provable Security - 8th International Conference, ProvSec 2014Document364 pagesProvable Security - 8th International Conference, ProvSec 2014alahbarPas encore d'évaluation
- General LPG Installation Guide PDFDocument60 pagesGeneral LPG Installation Guide PDFgheorghe garduPas encore d'évaluation
- Consumer Price SummaryDocument5 pagesConsumer Price SummaryKJ HiramotoPas encore d'évaluation
- Course Specifications: Fire Investigation and Failure Analysis (E901313)Document2 pagesCourse Specifications: Fire Investigation and Failure Analysis (E901313)danateoPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Wilson v. Baker Hughes Et. Al.Document10 pagesWilson v. Baker Hughes Et. Al.Patent LitigationPas encore d'évaluation
- Trading Book - AGDocument7 pagesTrading Book - AGAnilkumarGopinathanNairPas encore d'évaluation
- Anin, Cris Adrian U. Experiment Water Flirtation ELECTIVE 103Document2 pagesAnin, Cris Adrian U. Experiment Water Flirtation ELECTIVE 103Cris Adrian Umadac AninPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- SITXWHS001 - Participate in Safe Work Practices Student GuideDocument42 pagesSITXWHS001 - Participate in Safe Work Practices Student GuideMarianne FernandoPas encore d'évaluation
- Annual Presentation 18 19 EILDocument41 pagesAnnual Presentation 18 19 EILPartha Pratim GhoshPas encore d'évaluation
- Perrys Chemical Engineering Handbook 7th Edition PriceDocument3 pagesPerrys Chemical Engineering Handbook 7th Edition PriceSteven Valentius0% (12)
- Put Them Into A Big Bowl. Serve The Salad in Small Bowls. Squeeze Some Lemon Juice. Cut The Fruits Into Small Pieces. Wash The Fruits. Mix The FruitsDocument2 pagesPut Them Into A Big Bowl. Serve The Salad in Small Bowls. Squeeze Some Lemon Juice. Cut The Fruits Into Small Pieces. Wash The Fruits. Mix The FruitsNithya SweetiePas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Ojt HRMDocument7 pagesOjt HRMArlyn Joy NacinoPas encore d'évaluation
- PCI Secure Software Standard v1 - 0Document67 pagesPCI Secure Software Standard v1 - 0Antonio ClimaPas encore d'évaluation
- DS WhitePaper Troubleshooting 3DEXPERIENCE ABEND SituationsDocument26 pagesDS WhitePaper Troubleshooting 3DEXPERIENCE ABEND SituationsSam AntonyPas encore d'évaluation
- Zoomlion Gulf FZE Introduction: 1.1 ME Service Support 1.2 Construction CasesDocument13 pagesZoomlion Gulf FZE Introduction: 1.1 ME Service Support 1.2 Construction CasesArk TradingPas encore d'évaluation
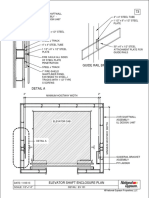
- Guide Rail Bracket AssemblyDocument1 pageGuide Rail Bracket AssemblyPrasanth VarrierPas encore d'évaluation
- Grammar and Oral Language Development (GOLD) : Reported By: Melyn A. Bacolcol Kate Batac Julie Ann OcampoDocument17 pagesGrammar and Oral Language Development (GOLD) : Reported By: Melyn A. Bacolcol Kate Batac Julie Ann Ocampoclara dupitasPas encore d'évaluation
- WP05 - ACT 01 - Development 1909Document53 pagesWP05 - ACT 01 - Development 1909ramesh9966Pas encore d'évaluation
- CHAPTER I KyleDocument13 pagesCHAPTER I KyleCresiel Pontijon100% (1)
- EverServ 7700 M77XX Quick Reference GuideDocument2 pagesEverServ 7700 M77XX Quick Reference GuidetangocharliepdxPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)