Vous aimerez peut-être aussi
- Formación de AnticuerposDocument19 pagesFormación de Anticuerposclarck777Pas encore d'évaluation
- Examen de FisiologiaDocument6 pagesExamen de FisiologiaGerman EduardoPas encore d'évaluation
- Biología Molecular y Celular de Los Tejidos Periodontales Sanos y Enfermos PDFDocument22 pagesBiología Molecular y Celular de Los Tejidos Periodontales Sanos y Enfermos PDFMafe TierradentroPas encore d'évaluation
- Anemia Megaloblastica - Medicina Clinica PDFDocument4 pagesAnemia Megaloblastica - Medicina Clinica PDFEbony Mariset Terrones DiazPas encore d'évaluation
- 6 Activacion de LinfocitosDocument52 pages6 Activacion de LinfocitosRocio GallegosPas encore d'évaluation
- Resumen InmunoglobulinasDocument20 pagesResumen InmunoglobulinasGabriel MalcherPas encore d'évaluation
- Sistema de Complemento y Sistema Inmune de Mucosas - InflamacionDocument32 pagesSistema de Complemento y Sistema Inmune de Mucosas - InflamacionSantiago CortesPas encore d'évaluation
- Criptococcus 2021Document26 pagesCriptococcus 2021Natashs Montalvo100% (1)
- Células y Moléculas Del Sistema Inmune InnatoDocument20 pagesCélulas y Moléculas Del Sistema Inmune InnatoFernando Aponte R.Pas encore d'évaluation
- Lectura de InmunidadDocument8 pagesLectura de InmunidadKatherine Milagros Apcho VeraPas encore d'évaluation
- Farmacos Antitusigenos FMH UNACHDocument26 pagesFarmacos Antitusigenos FMH UNACHBeto GomezPas encore d'évaluation
- 2 Leucocitos PDFDocument67 pages2 Leucocitos PDFKassandra MurilloPas encore d'évaluation
- Inmunologia en El EmbarazoDocument36 pagesInmunologia en El EmbarazoDioser AvilaPas encore d'évaluation
- InmunogDocument11 pagesInmunogcarosolanoPas encore d'évaluation
- Cap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesDocument10 pagesCap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesLinda Andreita Rosales Rosales0% (1)
- Celulas y Moleculas Del Sistema Inmune InnatoDocument13 pagesCelulas y Moleculas Del Sistema Inmune InnatofrancoPas encore d'évaluation
- Sistema de ComplementoDocument17 pagesSistema de ComplementoCarlosErazoPas encore d'évaluation
- Quimiocinas y Receptores para Quimiocinas - ExpoDocument8 pagesQuimiocinas y Receptores para Quimiocinas - ExpoJoseNeiserCastilloArrascuePas encore d'évaluation
- Mecanismo de Patogenicidad BacterianaDocument3 pagesMecanismo de Patogenicidad BacterianaKeyla Vente VelascoPas encore d'évaluation
- Toxoplasmosis y EmbarazoDocument16 pagesToxoplasmosis y EmbarazoBladimir ViloriaPas encore d'évaluation
- Protozoos PDFDocument213 pagesProtozoos PDFalexPas encore d'évaluation
- Cicatrizacion de Los TejidosDocument21 pagesCicatrizacion de Los TejidosLft Ediel PiñaPas encore d'évaluation
- Resumen MHCDocument3 pagesResumen MHCLuis GuzmánPas encore d'évaluation
- Resumen de LeucemiasDocument9 pagesResumen de LeucemiasDulce SalazarPas encore d'évaluation
- Linfocitos TDocument8 pagesLinfocitos TCristian Ignacio JaquePas encore d'évaluation
- Inmunidad Frente A Los TumoresDocument4 pagesInmunidad Frente A Los TumoresJacs SaulPas encore d'évaluation
- Respuesta Inmune HongosDocument12 pagesRespuesta Inmune HongosJesús Enrique Cantú BustosPas encore d'évaluation
- Activación Del Complemento Por IgGDocument13 pagesActivación Del Complemento Por IgGDr. HuijaPas encore d'évaluation
- 02 Células InmunocompetentesDocument10 pages02 Células InmunocompetentesIsa OcañaPas encore d'évaluation
- ClostridiumDocument42 pagesClostridiumAaron Villarreal PereyraPas encore d'évaluation
- Seminario de InvestigaciónDocument11 pagesSeminario de InvestigaciónAndres Felipe BotelloPas encore d'évaluation
- LisosomasDocument14 pagesLisosomasAndrea0% (1)
- Inmunodeficiencias Primarias y SecundariasDocument46 pagesInmunodeficiencias Primarias y Secundariasstephani hernandezPas encore d'évaluation
- CriptocococisDocument45 pagesCriptocococisMarcos Moreno100% (1)
- Histologia de Arterias y VenasDocument5 pagesHistologia de Arterias y Venassirley100% (2)
- Respuesta Adquirida (Inmunidad Adaptativa)Document17 pagesRespuesta Adquirida (Inmunidad Adaptativa)Orbely Garcia100% (1)
- Apoptosis Muerte Celular ProgramadaDocument7 pagesApoptosis Muerte Celular ProgramadaClaw JuárezPas encore d'évaluation
- Ciclo Celular Mitosis y Meiosis LecturaDocument24 pagesCiclo Celular Mitosis y Meiosis LecturaHaydee Ramirez LopezPas encore d'évaluation
- Bases Moleculares de La Diapedesis PDFDocument11 pagesBases Moleculares de La Diapedesis PDFRichard C. Munares100% (1)
- Reacción en Cadena de La PolimerasaDocument14 pagesReacción en Cadena de La PolimerasaZumba MkPas encore d'évaluation
- Autoinmunidad y TransplanteDocument49 pagesAutoinmunidad y TransplanteElia Salazar100% (1)
- Rechazo de TrasplanteDocument19 pagesRechazo de TrasplanteyoshaPas encore d'évaluation
- Tejido conectivo-2020-USMP PDFDocument54 pagesTejido conectivo-2020-USMP PDFSergio Contreras RomeroPas encore d'évaluation
- Clase 2 Generalidades de Los Hongos 2015Document43 pagesClase 2 Generalidades de Los Hongos 2015David Esteves Echanique100% (1)
- Cap 9. Órganos Linfoides y Ontogenia de Los LinfocitosDocument21 pagesCap 9. Órganos Linfoides y Ontogenia de Los LinfocitosAnthony Ramos FaríasPas encore d'évaluation
- Complejo Mayor de HistocompatibilidadDocument12 pagesComplejo Mayor de HistocompatibilidadElizabeth Travez ProañoPas encore d'évaluation
- Practica Sobre Medula OseaDocument6 pagesPractica Sobre Medula OseaCarlos CuencAPas encore d'évaluation
- 2.3 Inmunología de La LHDocument34 pages2.3 Inmunología de La LHVicente PalaciosPas encore d'évaluation
- Programa Respuesta Inmune 20-1Document3 pagesPrograma Respuesta Inmune 20-1Ruth Cobo RosalesPas encore d'évaluation
- Tema 9 MicobacteriasDocument72 pagesTema 9 MicobacteriasJesus VilchezPas encore d'évaluation
- Notas Borrador Sobre AlzheimerDocument4 pagesNotas Borrador Sobre AlzheimerSofíaPas encore d'évaluation
- Tejido MuscularDocument9 pagesTejido MuscularCristina Gómez RevueltaPas encore d'évaluation
- LeishmaniasisDocument7 pagesLeishmaniasisStefania BeltranPas encore d'évaluation
- Histología Del Sistema InmuneDocument9 pagesHistología Del Sistema InmuneJosé Luis Juárez VegaPas encore d'évaluation
- Deficiencia de G6P-DeshidrogenasaDocument49 pagesDeficiencia de G6P-DeshidrogenasaCésar AmcPas encore d'évaluation
- Farmacologia Capitulo 52 PDFDocument21 pagesFarmacologia Capitulo 52 PDFZuey Velasquez100% (1)
- Determinantes Sociales de La SaludDocument7 pagesDeterminantes Sociales de La SaludMónica CalderónPas encore d'évaluation
- Mecanismos de Defensa Del HuespedDocument61 pagesMecanismos de Defensa Del HuespedJessica Zegarra ArgumedoPas encore d'évaluation
- Purpura TrombocitopenicaDocument6 pagesPurpura TrombocitopenicaOlivia MaroPas encore d'évaluation
- Patogenicidad PDFDocument61 pagesPatogenicidad PDFLaboratoriosClinicosZaragozaPas encore d'évaluation
- Exposicion de EmbriologiaDocument33 pagesExposicion de EmbriologiaKaren HernandezPas encore d'évaluation
- TGF-Beta Información Complementaria Sobre La EstructuraDocument5 pagesTGF-Beta Información Complementaria Sobre La Estructurajohan.barreroPas encore d'évaluation
- Fase 1 - Construir Una Lista de Referencias Bibliográficas.Document4 pagesFase 1 - Construir Una Lista de Referencias Bibliográficas.paola villamarPas encore d'évaluation
- Diferencias Membrana Bacterias ArqueasDocument5 pagesDiferencias Membrana Bacterias ArqueasAndPas encore d'évaluation
- 6° - 5 Dominio Eukarya-Plantas y Animales II Periodo1Document2 pages6° - 5 Dominio Eukarya-Plantas y Animales II Periodo1Efrain Ducuara RamirezPas encore d'évaluation
- ContrabioticosDocument52 pagesContrabioticosIsmael Armando Collantes DiazPas encore d'évaluation
- Animales AcuáticosDocument17 pagesAnimales AcuáticosCathy LazcanoPas encore d'évaluation
- Taller BiocienciasDocument3 pagesTaller BiocienciasHugoHenaoMejia0% (1)
- Tarea 1 Individual BiodiversidadDocument1 pageTarea 1 Individual BiodiversidadAnalia CaramPas encore d'évaluation
- Que Es La Ingeniería GenéticaDocument14 pagesQue Es La Ingeniería GenéticaJHORDYN LESMESPas encore d'évaluation
- Biologia 02 BASES BIOLOGICASDocument9 pagesBiologia 02 BASES BIOLOGICASLeidy Cachique ShupingahuaPas encore d'évaluation
- Información Del Subgrupo 2 InmunologiaDocument10 pagesInformación Del Subgrupo 2 Inmunologiadiana ordoñezPas encore d'évaluation
- Biologia IntroDocument22 pagesBiologia IntroYudi Vilca PeñaPas encore d'évaluation
- Exp Arn SextoDocument25 pagesExp Arn Sextoruth escPas encore d'évaluation
- Biología en La SaludDocument9 pagesBiología en La SaludAna MataPas encore d'évaluation
- Induccion Fotoperiodica para Lograr Floracion en CDocument7 pagesInduccion Fotoperiodica para Lograr Floracion en CEdwin SosaPas encore d'évaluation
- Guia de Estudio Unidad I. Embriologia GeneralDocument48 pagesGuia de Estudio Unidad I. Embriologia GeneralMilagros GarciaPas encore d'évaluation
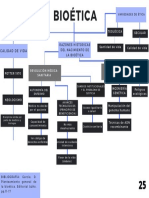
- Mapa Conceptual BioéticaDocument1 pageMapa Conceptual BioéticaAilyn LjPas encore d'évaluation
- Epigenética y DolorDocument27 pagesEpigenética y DolorPaloma Nxbzxnn100% (1)
- Diapositivas PtroteinaDocument29 pagesDiapositivas PtroteinarodrigoPas encore d'évaluation
- Reporte 6 JRQDocument5 pagesReporte 6 JRQJacqueline Rodríguez QuijanoPas encore d'évaluation
- Hipogonadismo - Incluye HH, KlinefelterDocument20 pagesHipogonadismo - Incluye HH, KlinefelterGabrielaPas encore d'évaluation
- Teorias Del EnvejecimientoDocument19 pagesTeorias Del Envejecimientoyakelin shakira100% (1)
- Biologia Celular e Histologia 1718 PDFDocument18 pagesBiologia Celular e Histologia 1718 PDFWall-e BurgosPas encore d'évaluation
- Bacterias y VirusDocument1 pageBacterias y VirusAldo Juarez100% (1)
- Genòmica-Trabajo de InvestigacionDocument8 pagesGenòmica-Trabajo de InvestigacionjessicPas encore d'évaluation
- AMINOACIDOSDocument4 pagesAMINOACIDOSANGEL ADRIAN ALTAMIRANO CONTRERASPas encore d'évaluation
- Troya - SJ - ResumenDocument3 pagesTroya - SJ - ResumenmariePas encore d'évaluation
- Resumen Bioantropología Tema IIDocument17 pagesResumen Bioantropología Tema IIEtienne DegreéfPas encore d'évaluation
- Fecundación y DesarrolloDocument23 pagesFecundación y DesarrolloMarcela TorreblancaPas encore d'évaluation