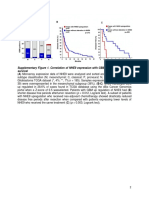
Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Acid Base Disorders JAPIDocument5 pagesAcid Base Disorders JAPIVitrag_Shah_1067Pas encore d'évaluation
- Aamc 4RDocument76 pagesAamc 4RBellony Sanders100% (2)
- JC-2 Tutorial-1 Immunology Practice MCQ'sDocument14 pagesJC-2 Tutorial-1 Immunology Practice MCQ'shelamahjoubmounirdmo100% (5)
- Ironfang Invasion - Players GuideDocument12 pagesIronfang Invasion - Players GuideThomas McDonaldPas encore d'évaluation
- BTS Sarcoidosis Clinical StatementDocument17 pagesBTS Sarcoidosis Clinical StatementDarrin WiebePas encore d'évaluation
- Oet Test Material: Occupational English Test Reading Sub-Test NursingDocument13 pagesOet Test Material: Occupational English Test Reading Sub-Test NursingRyu Tse33% (3)
- Prioritization - FNCPDocument10 pagesPrioritization - FNCPJeffer Dancel67% (3)
- Clinical Practice Guidelines - 2017Document54 pagesClinical Practice Guidelines - 2017Cem ÜnsalPas encore d'évaluation
- Cellular Aberration Acute Biologic Crisis 100 Items - EditedDocument8 pagesCellular Aberration Acute Biologic Crisis 100 Items - EditedSherlyn PedidaPas encore d'évaluation
- Chapter 15 Acid Base Equilibrium and BuffersDocument9 pagesChapter 15 Acid Base Equilibrium and Buffersmememe123123Pas encore d'évaluation
- Summary of MetabolismDocument33 pagesSummary of Metabolismmememe123123Pas encore d'évaluation
- Master AAMC MCAT-2015 Topics List: Reorganized and Duplicates Removed 1. Amino AcidsDocument30 pagesMaster AAMC MCAT-2015 Topics List: Reorganized and Duplicates Removed 1. Amino AcidsSukhvir AujlaPas encore d'évaluation
- Ochem 5 Lab TechniquesDocument50 pagesOchem 5 Lab Techniquesmememe123123Pas encore d'évaluation
- Khan Academy Notes - CellsDocument42 pagesKhan Academy Notes - Cellsmememe123123100% (1)
- Hi Colt, I Would Like To Follow Up With You in Regards To The Seat Cushion Frame Support Wire. Thank YouDocument1 pageHi Colt, I Would Like To Follow Up With You in Regards To The Seat Cushion Frame Support Wire. Thank Youmememe123123Pas encore d'évaluation
- Leitz & Kavalali 2011Document9 pagesLeitz & Kavalali 2011mememe123123Pas encore d'évaluation
- PNAS 2003 Al Hajj 3983 8Document8 pagesPNAS 2003 Al Hajj 3983 8mememe123123Pas encore d'évaluation
- Topic8Assignment CritiqueArticleDocument5 pagesTopic8Assignment CritiqueArticlearhcee11Pas encore d'évaluation
- Cancer Bio Exam Article Supplemental InfoDocument8 pagesCancer Bio Exam Article Supplemental Infomememe123123Pas encore d'évaluation
- Microbial GeneticsDocument11 pagesMicrobial GeneticsJennie M. MagdirilaPas encore d'évaluation
- What's On The MCAT 2015Document128 pagesWhat's On The MCAT 2015kape1onePas encore d'évaluation
- 10 Alkenes ReactionsDocument1 page10 Alkenes ReactionsRakesh Kumar SutharPas encore d'évaluation
- Master AAMC MCAT-2015 Topics List: Reorganized and Duplicates Removed 1. Amino AcidsDocument30 pagesMaster AAMC MCAT-2015 Topics List: Reorganized and Duplicates Removed 1. Amino AcidsSukhvir AujlaPas encore d'évaluation
- 10 Alkenes ReactionsDocument1 page10 Alkenes ReactionsRakesh Kumar SutharPas encore d'évaluation
- C242f14sec1syllabus PDFDocument20 pagesC242f14sec1syllabus PDFmememe123123Pas encore d'évaluation
- MCAT Study PlanDocument55 pagesMCAT Study Planmememe123123Pas encore d'évaluation
- Chem 242 Presentation SummaryDocument2 pagesChem 242 Presentation Summarymememe123123Pas encore d'évaluation
- MCAT Khan Academy NotesDocument1 pageMCAT Khan Academy Notesmememe123123Pas encore d'évaluation
- C 242 F 14 T 1Document15 pagesC 242 F 14 T 1mememe123123Pas encore d'évaluation
- MCAT 2015 OutlineDocument128 pagesMCAT 2015 OutlinequirkyheadbandsPas encore d'évaluation
- Reference FormDocument2 pagesReference Formmememe123123Pas encore d'évaluation
- Malawi Clinical HIV Guidelines 2019 Addendumversion 8.1Document28 pagesMalawi Clinical HIV Guidelines 2019 Addendumversion 8.1INNOCENT KHULIWAPas encore d'évaluation
- Fertilization Reflection PaperDocument2 pagesFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- Sample ReportDocument3 pagesSample ReportRobeants Charles PierrePas encore d'évaluation
- Growth Comparison in Children With and Without Food Allergies in 2 Different Demographic PopulationsDocument7 pagesGrowth Comparison in Children With and Without Food Allergies in 2 Different Demographic PopulationsMaria Agustina Sulistyo WulandariPas encore d'évaluation
- GENERAL RISK ASSESSMENT Mechatronics LaboratoryDocument2 pagesGENERAL RISK ASSESSMENT Mechatronics LaboratoryJason TravisPas encore d'évaluation
- Test Container Form Lab Process Remarks HKL (Hospital Appointment Date - Specialist's SignatureDocument9 pagesTest Container Form Lab Process Remarks HKL (Hospital Appointment Date - Specialist's SignaturesyasyaPas encore d'évaluation
- Neurobiology of Sleep: Madhu Kalia4Document5 pagesNeurobiology of Sleep: Madhu Kalia4Julian ReyesPas encore d'évaluation
- Clinchers: RheumatologyDocument22 pagesClinchers: RheumatologyvarrakeshPas encore d'évaluation
- ZFN, TALEN, and CRISPR-Cas-based Methods For Genome EngineeringDocument9 pagesZFN, TALEN, and CRISPR-Cas-based Methods For Genome EngineeringRomina Tamara Gil RamirezPas encore d'évaluation
- International Rice Research Notes Vol.18 No.1Document69 pagesInternational Rice Research Notes Vol.18 No.1ccquintosPas encore d'évaluation
- Autism Research Paper OutlineDocument5 pagesAutism Research Paper Outlineafmcifezi100% (1)
- Journals With No Article Processing ChargesDocument14 pagesJournals With No Article Processing ChargesNurul Jami Bakti JayaPas encore d'évaluation
- Polydactyly of The Foot A Review.92Document10 pagesPolydactyly of The Foot A Review.92mamyeu1801Pas encore d'évaluation
- Cancer BulletsDocument3 pagesCancer Bulletsraquel maniegoPas encore d'évaluation
- Arthur CrawfordDocument308 pagesArthur CrawfordRupali Mokashi100% (1)
- Punjab Municipal Corporation Act, 1976 PDFDocument180 pagesPunjab Municipal Corporation Act, 1976 PDFSci UpscPas encore d'évaluation
- ESC - 2021 - The Growing Role of Genetics in The Understanding of Cardiovascular Diseases - Towards Personalized MedicineDocument5 pagesESC - 2021 - The Growing Role of Genetics in The Understanding of Cardiovascular Diseases - Towards Personalized MedicineDini SuhardiniPas encore d'évaluation
- University of Cambridge International Examinations General Certificate of Education Ordinary LevelDocument16 pagesUniversity of Cambridge International Examinations General Certificate of Education Ordinary Levelmstudy123456Pas encore d'évaluation
- AUB - Microscopic Analysis of UrineDocument4 pagesAUB - Microscopic Analysis of UrineJeanne Rodiño100% (1)
- Beauty Hankering For Memory: Marne L. KilatesDocument9 pagesBeauty Hankering For Memory: Marne L. KilatesKuya ReaGoPas encore d'évaluation
- FOCGB3 Utest DLR 6BDocument2 pagesFOCGB3 Utest DLR 6BGyurácz GyulaPas encore d'évaluation
- Biology Viral DiseasesDocument11 pagesBiology Viral DiseasesPrasoon Singh RajputPas encore d'évaluation