Vous aimerez peut-être aussi
- Iaquinto 2010Document6 pagesIaquinto 2010Altair LopesPas encore d'évaluation
- The Complete Guide To Food For Sports Performance - Peak Nutrition For Your Sport (Team Nanban) TMRGDocument545 pagesThe Complete Guide To Food For Sports Performance - Peak Nutrition For Your Sport (Team Nanban) TMRGdnaskar92% (13)
- 2017 Journal Citation Reports by Thomson Reuters: July 2017Document251 pages2017 Journal Citation Reports by Thomson Reuters: July 2017vlad lupasteanuPas encore d'évaluation
- Cscca and Nsca Joint Consensus Guidelines For Transition Periods: Safe Return To Training Following InactivityDocument23 pagesCscca and Nsca Joint Consensus Guidelines For Transition Periods: Safe Return To Training Following InactivityAltair LopesPas encore d'évaluation
- Personal Trainer Pocketbook - A Handy Reference For All Your Daily Questions (PDFDrive) PDFDocument136 pagesPersonal Trainer Pocketbook - A Handy Reference For All Your Daily Questions (PDFDrive) PDFAakash KoulPas encore d'évaluation
- The Effects of Time of Day-Specific Resistance Training On Adaptations in Skeletal Muscle Hypertrophy and Muscle Strength: A Systematic Review and Meta-AnalysisDocument13 pagesThe Effects of Time of Day-Specific Resistance Training On Adaptations in Skeletal Muscle Hypertrophy and Muscle Strength: A Systematic Review and Meta-AnalysisAltair LopesPas encore d'évaluation
- Escala de Status de RecuperacaoDocument10 pagesEscala de Status de RecuperacaoAltair LopesPas encore d'évaluation
- Charles Glass - The Fundamentals of Bodybuilding and Physique Sculpting PDFDocument35 pagesCharles Glass - The Fundamentals of Bodybuilding and Physique Sculpting PDFAkhilesh Sankar92% (49)
- The Effect of Schroth Therapy On Thoracic KyphoticDocument10 pagesThe Effect of Schroth Therapy On Thoracic KyphoticAltair LopesPas encore d'évaluation
- Lee 2010 HJKDocument12 pagesLee 2010 HJKAltair LopesPas encore d'évaluation
- Damas 2016Document27 pagesDamas 2016Altair LopesPas encore d'évaluation
- Partial Vs Full Range Repetition For HypertrophyDocument13 pagesPartial Vs Full Range Repetition For HypertrophyPrecision Wellness Precision WellnessPas encore d'évaluation
- Exercise For Hypertension: A Prescription Update Integrating Existing Recommendations With Emerging ResearchDocument10 pagesExercise For Hypertension: A Prescription Update Integrating Existing Recommendations With Emerging ResearchNuridhaLatifPas encore d'évaluation
- Ginástica Laboral QuestionárioDocument2 pagesGinástica Laboral QuestionárioAltair LopesPas encore d'évaluation
- CXCXXCDocument6 pagesCXCXXCAltair LopesPas encore d'évaluation
- Training-Induced Changes in Muscle Architecture and Specific TensionDocument7 pagesTraining-Induced Changes in Muscle Architecture and Specific TensionAltair LopesPas encore d'évaluation
- Training-Induced Changes in Muscle Architecture and Specific TensionDocument7 pagesTraining-Induced Changes in Muscle Architecture and Specific TensionAltair LopesPas encore d'évaluation
- Journal of Sports SciencesDocument8 pagesJournal of Sports SciencesAltair LopesPas encore d'évaluation
- Cellular and Molecular Mechanisms of MuscleDocument15 pagesCellular and Molecular Mechanisms of MuscleAltair LopesPas encore d'évaluation
- 17825Document5 pages17825Altair LopesPas encore d'évaluation
- 17828Document6 pages17828Altair LopesPas encore d'évaluation
- PDFDocument6 pagesPDFAltair LopesPas encore d'évaluation
- Kettlebell Swing and Lumbar LoadsDocument12 pagesKettlebell Swing and Lumbar Loadsrearickstrength_comPas encore d'évaluation
- 17828Document6 pages17828Altair LopesPas encore d'évaluation
- 3846123Document7 pages3846123Altair LopesPas encore d'évaluation
- WNL 0000000000000551 PDFDocument9 pagesWNL 0000000000000551 PDFAltair LopesPas encore d'évaluation
- 17826Document5 pages17826Altair LopesPas encore d'évaluation
- GHGHDocument16 pagesGHGHAltair LopesPas encore d'évaluation
- Muscle & Nerve Volume 26 Issue 5 2002 (Doi 10.1002/mus.10225)Document5 pagesMuscle & Nerve Volume 26 Issue 5 2002 (Doi 10.1002/mus.10225)Altair LopesPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Board Review Endocrinology A. ApiradeeDocument47 pagesBoard Review Endocrinology A. ApiradeePiyasak NaumnaPas encore d'évaluation
- The Impact of StressDocument3 pagesThe Impact of StressACabalIronedKryptonPas encore d'évaluation
- Tumors of The Central Nervous System - VOL 12Document412 pagesTumors of The Central Nervous System - VOL 12vitoPas encore d'évaluation
- Dip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Document1 pageDip Obst (SA) Past Papers - 2020 1st Semester 1-6-2023Neo Latoya MadunaPas encore d'évaluation
- GTT Module 5Document156 pagesGTT Module 5ABDULRAHIMAN RAJEKHANPas encore d'évaluation
- 3-Step Mindset Reset: Overcome Self-Doubt with Mel Robbins' TrainingDocument11 pages3-Step Mindset Reset: Overcome Self-Doubt with Mel Robbins' TrainingBožana RadošPas encore d'évaluation
- NCMA 217 - Newborn Assessment Ma'am JhalDocument5 pagesNCMA 217 - Newborn Assessment Ma'am JhalMariah Blez BognotPas encore d'évaluation
- Chapter 3 - CT&VT - Part 1Document63 pagesChapter 3 - CT&VT - Part 1zhafran100% (1)
- Micdak BackgroundDocument3 pagesMicdak Backgroundappiah ernestPas encore d'évaluation
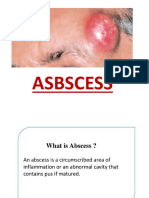
- ABSCESSDocument35 pagesABSCESSlax prajapatiPas encore d'évaluation
- FinalsDocument8 pagesFinalsDumpPas encore d'évaluation
- Proper restraint techniques for dogs and catsDocument153 pagesProper restraint techniques for dogs and catsjademattican75% (4)
- Live Performance Award Ma000081 Pay GuideDocument48 pagesLive Performance Award Ma000081 Pay GuideDan LijndersPas encore d'évaluation
- Challenges of Tourism Students in Commercial CookingDocument29 pagesChallenges of Tourism Students in Commercial CookingMaeve Anne JaimePas encore d'évaluation
- English III Module 2 Simple Present Job and Job VerbsDocument4 pagesEnglish III Module 2 Simple Present Job and Job VerbsAdrian CortesPas encore d'évaluation
- BCM Continuous ImprovementDocument22 pagesBCM Continuous ImprovementnikoPas encore d'évaluation
- RTG E-One - Manual de Manutenção 41300-41303 (EN)Document328 pagesRTG E-One - Manual de Manutenção 41300-41303 (EN)Conrado Soares100% (1)
- Endocrown Review 1Document9 pagesEndocrown Review 1Anjali SatsangiPas encore d'évaluation
- A&P 2 - Digestive System Flashcards - QuizletDocument1 pageA&P 2 - Digestive System Flashcards - QuizletMunachande KanondoPas encore d'évaluation
- Solution Manual of Physics by Arthur BeiserDocument145 pagesSolution Manual of Physics by Arthur BeiserManuull71% (49)
- Grade Eleven Test 2019 Social StudiesDocument6 pagesGrade Eleven Test 2019 Social StudiesClair VickeriePas encore d'évaluation
- Soal Upk B Inggris PKBM WinaDocument11 pagesSoal Upk B Inggris PKBM WinaCuman MitosPas encore d'évaluation
- Q4 Module 8Document14 pagesQ4 Module 8DerickPas encore d'évaluation
- Formularium ApotekDocument12 pagesFormularium ApotekNurul Evi kurniatiPas encore d'évaluation
- Li Ching Wing V Xuan Yi Xiong (2004) 1 HKC 353Document11 pagesLi Ching Wing V Xuan Yi Xiong (2004) 1 HKC 353hPas encore d'évaluation
- Elem. Reading PracticeDocument10 pagesElem. Reading PracticeElissa Janquil RussellPas encore d'évaluation
- General Specifications: Detail ADocument1 pageGeneral Specifications: Detail AJeniel PascualPas encore d'évaluation
- Consumer Behaviour: Group ProjectDocument5 pagesConsumer Behaviour: Group ProjectAanchal MahajanPas encore d'évaluation
- Junayed - 19 39800 1Document11 pagesJunayed - 19 39800 1gurujeePas encore d'évaluation
- Emission of Volatile Organic Compounds (Vocs) From Dispersion and Cementitious Waterproofing ProductsDocument16 pagesEmission of Volatile Organic Compounds (Vocs) From Dispersion and Cementitious Waterproofing ProductsKrishna KusumaPas encore d'évaluation