Vous aimerez peut-être aussi
- Tincion H-EDocument3 pagesTincion H-EDavide ManciniPas encore d'évaluation
- Civil PDFDocument11 pagesCivil PDFDavide ManciniPas encore d'évaluation
- Curriculum Vitae - Doc Nueva Imagen de CirrrDocument2 pagesCurriculum Vitae - Doc Nueva Imagen de CirrrDavide ManciniPas encore d'évaluation
- Higado 1Document23 pagesHigado 1Davide ManciniPas encore d'évaluation
- Banco Preguntas QuimicaDocument94 pagesBanco Preguntas QuimicaMario Gonzalo Henriquez PardoPas encore d'évaluation
- Tejido ÓseoDocument4 pagesTejido ÓseoDavide ManciniPas encore d'évaluation
- La Placenta, Estructura y FuncionDocument3 pagesLa Placenta, Estructura y FuncionDavide ManciniPas encore d'évaluation
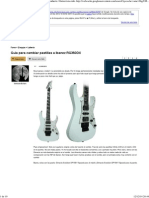
- Guia Pastillas GuitarraDocument10 pagesGuia Pastillas GuitarraMartinFebreiroPas encore d'évaluation
- Caso InditexDocument13 pagesCaso Inditexelvis2709487100% (2)
- Conclusiones Matriz de Analisis de Riesgos (Grupo)Document6 pagesConclusiones Matriz de Analisis de Riesgos (Grupo)Suzette Bowie100% (1)
- Senos Magnificados PDF, Libro de Liz CabreraDocument18 pagesSenos Magnificados PDF, Libro de Liz CabreraAntonio VallejosPas encore d'évaluation
- Ats Trabajo Seguro Cosedora de SacosDocument3 pagesAts Trabajo Seguro Cosedora de SacosLaura MejiaPas encore d'évaluation
- Ocopilla Analisis CrecimientoDocument14 pagesOcopilla Analisis CrecimientoGabriel LeoPas encore d'évaluation
- Enfoque Sistemico, Una Introduccion A La Psicoterapia FamiliarDocument6 pagesEnfoque Sistemico, Una Introduccion A La Psicoterapia FamiliarBerenice De LeonPas encore d'évaluation
- Lo Que Se Aprende en Los Mejores MBA2008Document53 pagesLo Que Se Aprende en Los Mejores MBA2008PedroPas encore d'évaluation
- Ley TransitoriaaDocument6 pagesLey TransitoriaaAnita EscalantePas encore d'évaluation
- Propuesta Proyecto de PalmaDocument7 pagesPropuesta Proyecto de PalmaJonathan LaguadoPas encore d'évaluation
- Udi 7 8-2 y 8-3Document48 pagesUdi 7 8-2 y 8-3PEPELERIAPas encore d'évaluation
- T LBC 140280Document105 pagesT LBC 140280SergioArmandoGtzOrantesPas encore d'évaluation
- Topic 5 - MetalesDocument9 pagesTopic 5 - MetaleslunademarinaPas encore d'évaluation
- Mol530 s1 Estrategia Competitiva PorterDocument30 pagesMol530 s1 Estrategia Competitiva PorterJosselin Aguirre GarnicaPas encore d'évaluation
- 1ra Unidad Didactica - 2Document80 pages1ra Unidad Didactica - 2ELIO EDISON AMAO SACAPas encore d'évaluation
- Informe Diagrama Sinoptico Del ProcesoDocument5 pagesInforme Diagrama Sinoptico Del ProcesoRosa mariaPas encore d'évaluation
- Destreza 1Document3 pagesDestreza 1Fernando CuaranPas encore d'évaluation
- Evolucion Cientifica Del Estudio de La Microscopia y MicrometriaDocument16 pagesEvolucion Cientifica Del Estudio de La Microscopia y MicrometriaANGHELO JEANPIERRE MARTINEZ ELIZALDEPas encore d'évaluation
- Evaluacion PublisherDocument2 pagesEvaluacion PublisherMarielis anaya prasca100% (1)
- Trabajo Practico de Cinematica Fisica 3er TrimestreDocument5 pagesTrabajo Practico de Cinematica Fisica 3er TrimestreAnonymous ByHuesPas encore d'évaluation
- FMQ-006 Reporte Prueba de Presión HidrostáticaDocument1 pageFMQ-006 Reporte Prueba de Presión HidrostáticaAntonioPas encore d'évaluation
- Informe 5 RevenidoDocument11 pagesInforme 5 RevenidoJset EstevezPas encore d'évaluation
- 28 El Verdadero Sentido de La NavidadDocument1 page28 El Verdadero Sentido de La Navidadlizcornejorjm100% (1)
- Práctica 9 - Filtración Continua en Un Filtro de Tambor Rotatorio - 20131363Document27 pagesPráctica 9 - Filtración Continua en Un Filtro de Tambor Rotatorio - 20131363Arlahe Salazar Luna-0086Pas encore d'évaluation
- Tesis CimentacionesDocument178 pagesTesis CimentacionesMishel FuentesPas encore d'évaluation
- Acta de Conciliacion ExtrajudicialDocument2 pagesActa de Conciliacion ExtrajudicialAndres Camilo Bernal CamargoPas encore d'évaluation
- Estadistica Básica - Actividad SuplementariaDocument5 pagesEstadistica Básica - Actividad SuplementariaIker CastilloPas encore d'évaluation
- T2 Transporte y Distribucion.Document6 pagesT2 Transporte y Distribucion.luis javier de la cruz victorioPas encore d'évaluation
- Foro N 01Document5 pagesForo N 01Eduardo Paz castilloPas encore d'évaluation
- Información General AmtDocument4 pagesInformación General AmtPABLO GRANJAPas encore d'évaluation