Vous aimerez peut-être aussi
- Oxidative StressDocument53 pagesOxidative Stressapi-3723327100% (2)
- Lecture #10. The Biology of Cancer, p53Document49 pagesLecture #10. The Biology of Cancer, p53cafemedPas encore d'évaluation
- Herbs and SpicesDocument77 pagesHerbs and SpicesNicole RicohermosoPas encore d'évaluation
- Simple EmergenciesDocument177 pagesSimple Emergenciesharsh89100% (1)
- Spirulina Platensis Food For Future A Re 1Document8 pagesSpirulina Platensis Food For Future A Re 1SabrinaPas encore d'évaluation
- Thiamine: Group MembersDocument48 pagesThiamine: Group MembersJai-Michael FrancisPas encore d'évaluation
- The Human Gut MicrobiomeDocument11 pagesThe Human Gut MicrobiomeRenata RisiPas encore d'évaluation
- D8045-16 Standard Test Method For Acid Number of Crude Oils and Petroleum Products by CatalyticDocument6 pagesD8045-16 Standard Test Method For Acid Number of Crude Oils and Petroleum Products by CatalytichishamPas encore d'évaluation
- Head and NeckDocument11 pagesHead and NeckdrsamnPas encore d'évaluation
- HEALTH BENEFITS of Eucheuma CottoniiDocument3 pagesHEALTH BENEFITS of Eucheuma Cottoniije moguriPas encore d'évaluation
- Types of ProcurementDocument7 pagesTypes of ProcurementrahulPas encore d'évaluation
- Mineral NutritionDocument13 pagesMineral NutritionAashishPas encore d'évaluation
- Mycorrhizae: Plant Roots and FungiDocument29 pagesMycorrhizae: Plant Roots and FungigitaPas encore d'évaluation
- Alternative Medicine Therapies, Cures and Treatments: Vibratory Energy MedicineDocument9 pagesAlternative Medicine Therapies, Cures and Treatments: Vibratory Energy Medicinestereo1073Pas encore d'évaluation
- MD Anderson Cancer Center Report On Trametes Versicolor Turkey Tail MushroomDocument14 pagesMD Anderson Cancer Center Report On Trametes Versicolor Turkey Tail MushroomMichaelPas encore d'évaluation
- ISKCON Desire Tree - Brahma Vimohana LeelaDocument34 pagesISKCON Desire Tree - Brahma Vimohana LeelaISKCON desire treePas encore d'évaluation
- Advances in Mass Production Technology of Arbuscular MycorrhizaDocument13 pagesAdvances in Mass Production Technology of Arbuscular Mycorrhizarahulsinghrs100% (1)
- Fractal Neurodyamics and Quantum Chaos: Resolving The Mind-Brain Paradox Through Novel BiophysicsDocument24 pagesFractal Neurodyamics and Quantum Chaos: Resolving The Mind-Brain Paradox Through Novel Biophysicskhanverse100% (1)
- British Journal of Nutrition (2008), 99, E-Suppl.Document50 pagesBritish Journal of Nutrition (2008), 99, E-Suppl.nihadtoussounPas encore d'évaluation
- Preventing and Reversing Cancer - The Absolute Health Integrative Oncology MethodDocument56 pagesPreventing and Reversing Cancer - The Absolute Health Integrative Oncology MethodHalal Media MalaysiaPas encore d'évaluation
- Reusi Dat Ton The Origin of Nuat Boran by Danko Lara RadicDocument8 pagesReusi Dat Ton The Origin of Nuat Boran by Danko Lara RadicJacklynlim LkcPas encore d'évaluation
- Early Thiamine DeficiencyDocument175 pagesEarly Thiamine DeficiencyAndrea SirahPas encore d'évaluation
- Mechanism of the Anticancer Effect of PhytochemicalsD'EverandMechanism of the Anticancer Effect of PhytochemicalsPas encore d'évaluation
- The Human Microbiome at The Interface of Health and Disease PDFDocument11 pagesThe Human Microbiome at The Interface of Health and Disease PDFabcder1234Pas encore d'évaluation
- The Human Microbiome: D H U TADocument19 pagesThe Human Microbiome: D H U TADetox healing carePas encore d'évaluation
- Fan Tool Kit - Ad Hoc Group - V4ddDocument121 pagesFan Tool Kit - Ad Hoc Group - V4ddhmaza shakeelPas encore d'évaluation
- Medical Electricity,!: Dr. A. H. StevejstsDocument20 pagesMedical Electricity,!: Dr. A. H. Stevejstsaryaa2020Pas encore d'évaluation
- Plante BatteriesDocument16 pagesPlante Batteriessureshnfcl0% (1)
- MRCS1 2brochureDocument12 pagesMRCS1 2brochuredrsamn100% (2)
- Ciclogenia 1Document1 pageCiclogenia 1Rafael DurantePas encore d'évaluation
- Antimicrobial Properties of Coconut OilDocument5 pagesAntimicrobial Properties of Coconut OilIggystratPas encore d'évaluation
- The Prime Cause and Prevention of Disease WarburgDocument3 pagesThe Prime Cause and Prevention of Disease WarburgMichael DietrickPas encore d'évaluation
- r4f Pastest MnemonicsDocument23 pagesr4f Pastest MnemonicsplayuhzPas encore d'évaluation
- Acute Respiratory FailureDocument8 pagesAcute Respiratory FailureCayunk NorlianaPas encore d'évaluation
- In Vitro Culture of Orchids: The Roles of Class-1 Knox Gene in Shoot DevelopmentDocument10 pagesIn Vitro Culture of Orchids: The Roles of Class-1 Knox Gene in Shoot DevelopmentjammesPas encore d'évaluation
- Journal of Histochemistry & Cytochemistry: Reduction of Lipofuscin-Like Autofluorescence in Fluorescently Labeled TissueDocument13 pagesJournal of Histochemistry & Cytochemistry: Reduction of Lipofuscin-Like Autofluorescence in Fluorescently Labeled TissueLovadi Emese IldikóPas encore d'évaluation
- Bars Performance AppraisalDocument6 pagesBars Performance AppraisalPhillip Miler0% (1)
- FBS Q1 WK1Document4 pagesFBS Q1 WK1Nicole Eve Pelaez-AbarrientosPas encore d'évaluation
- Hookah Bar Business PlanDocument34 pagesHookah Bar Business PlanAbdelkebir LabyadPas encore d'évaluation
- Stress & HippocampusDocument26 pagesStress & HippocampusGEETA MOHANPas encore d'évaluation
- DLS Dynamic-Light-Scattering Teoria PDFDocument13 pagesDLS Dynamic-Light-Scattering Teoria PDFTatianaPas encore d'évaluation
- A Comprehensive Review On Watermelon Seed OilDocument7 pagesA Comprehensive Review On Watermelon Seed OilAndxp51Pas encore d'évaluation
- Medicinal Properties of Moringa Oleifera PDFDocument7 pagesMedicinal Properties of Moringa Oleifera PDFsufijinnPas encore d'évaluation
- Natural Farming PDFDocument2 pagesNatural Farming PDFGuruva ReddyPas encore d'évaluation
- Liposomal Drug DeliveryDocument13 pagesLiposomal Drug DeliveryLuisMaldonadoPas encore d'évaluation
- The Effect of Different Light Conditions On The Growth PDFDocument22 pagesThe Effect of Different Light Conditions On The Growth PDFSciResearchPas encore d'évaluation
- Controlling Ticks and Tick-Borne DiseasesDocument12 pagesControlling Ticks and Tick-Borne Diseasesapi-3703629100% (1)
- Mushroom MagazineDocument7 pagesMushroom MagazineyukichanPas encore d'évaluation
- NaCl: Silver NanoparticlesDocument12 pagesNaCl: Silver NanoparticlesWhy BotherPas encore d'évaluation
- Unesco - Eolss Sample Chapter: Plants As A Source of Anti-Cancer AgentsDocument15 pagesUnesco - Eolss Sample Chapter: Plants As A Source of Anti-Cancer AgentsSundararajan Jeyaraman100% (1)
- Oregano Benefits 2beingfitDocument8 pagesOregano Benefits 2beingfitK AnjaliPas encore d'évaluation
- Targeting Mitochondria For Cancer TherapyDocument18 pagesTargeting Mitochondria For Cancer Therapysatya_chagantiPas encore d'évaluation
- Common Themes 1Document6 pagesCommon Themes 1api-356397852Pas encore d'évaluation
- IRPS 138 The Genus Oryza L. Current Status of TaxonomyDocument22 pagesIRPS 138 The Genus Oryza L. Current Status of TaxonomyIRRI_resourcesPas encore d'évaluation
- Olympus Imt 2 Instructions PDFDocument28 pagesOlympus Imt 2 Instructions PDFOpeyemi AjayiPas encore d'évaluation
- PRIME PubMed - A Bioavailability Study Comparing Two Oral Formulations Containing Zinc (ZN Bis-Glycinate vs. ZN Gluconate) After A Single Administration To Twelve Healthy Female Volunteers PDFDocument2 pagesPRIME PubMed - A Bioavailability Study Comparing Two Oral Formulations Containing Zinc (ZN Bis-Glycinate vs. ZN Gluconate) After A Single Administration To Twelve Healthy Female Volunteers PDFYacine BoumetlouaPas encore d'évaluation
- Toxicology For Activists: Romeo F. Quijano, M.DDocument87 pagesToxicology For Activists: Romeo F. Quijano, M.DRomeo QuijanoPas encore d'évaluation
- 17april2008 Brix PresentationDocument10 pages17april2008 Brix PresentationDilannirangaPas encore d'évaluation
- Natural Antiviral PlantDocument19 pagesNatural Antiviral PlantGaby OliveraPas encore d'évaluation
- Plant Responses To Flooding Stress PDFDocument8 pagesPlant Responses To Flooding Stress PDFTica DuyênPas encore d'évaluation
- Pharmacognosy-I (Part-3)Document39 pagesPharmacognosy-I (Part-3)LibAmauPas encore d'évaluation
- Psoriasis - Pathophysiology, Conventional, and Alternative Approaches To TreatmentDocument24 pagesPsoriasis - Pathophysiology, Conventional, and Alternative Approaches To TreatmentGede Eka Putra NugrahaPas encore d'évaluation
- Goat Under CoconutDocument3 pagesGoat Under Coconutjhoe eliasPas encore d'évaluation
- HookwormDocument9 pagesHookwormfadilah mutiaPas encore d'évaluation
- Capsaicinoids - Pungency Beyond Capsicum - E R Naves Et Al - Tplants 2018Document12 pagesCapsaicinoids - Pungency Beyond Capsicum - E R Naves Et Al - Tplants 2018notoPas encore d'évaluation
- Medicine - IJMPS - Black Cumin - Nigella Sativa L. - Affects - Halla Muhee NajiDocument10 pagesMedicine - IJMPS - Black Cumin - Nigella Sativa L. - Affects - Halla Muhee NajiTJPRC PublicationsPas encore d'évaluation
- Biophotons As Neural Communication Signals PDFDocument8 pagesBiophotons As Neural Communication Signals PDFweb3351Pas encore d'évaluation
- Antibacterial Activity of Propolis Produced by Trigona Spp. Against Campylobacter SPPDocument4 pagesAntibacterial Activity of Propolis Produced by Trigona Spp. Against Campylobacter SPPAmin FatoniPas encore d'évaluation
- Rebuilding Human Ability To Regenerate Cells Inhibiting Anatomical Growth During Early Embryonic StageDocument8 pagesRebuilding Human Ability To Regenerate Cells Inhibiting Anatomical Growth During Early Embryonic StageInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- Stem Cells and Repair of Lung InjuriesDocument9 pagesStem Cells and Repair of Lung InjuriesDani PazPas encore d'évaluation
- Stem Cells Past, Present and FutureDocument3 pagesStem Cells Past, Present and FutureAndrés ChávezPas encore d'évaluation
- Range of Vertebrate Regenerative Tissues: Regeneration 1&2Document77 pagesRange of Vertebrate Regenerative Tissues: Regeneration 1&2AsimMohammad-NaseerPas encore d'évaluation
- Sciadv Aao4881Document11 pagesSciadv Aao4881randomaeiou7273Pas encore d'évaluation
- Biomaterial and Stem Cell-Based Strategies For Skeletal Muscle RegenerationDocument17 pagesBiomaterial and Stem Cell-Based Strategies For Skeletal Muscle RegenerationAntonio JuniorPas encore d'évaluation
- Public Writing Band DescriptorsDocument2 pagesPublic Writing Band DescriptorsKaveh JalaliPas encore d'évaluation
- Heart FailureDocument10 pagesHeart FailuredrsamnPas encore d'évaluation
- IELTS Speaking Criteria Public Version PDFDocument1 pageIELTS Speaking Criteria Public Version PDFDiana Laura Durán Chablé100% (1)
- Articles PracticalDocument4 pagesArticles PracticaldrsamnPas encore d'évaluation
- Approaches To Management of Acute Pain 1Document22 pagesApproaches To Management of Acute Pain 1drsamnPas encore d'évaluation
- Oxygen Therapy and Oxygen ToxicityDocument8 pagesOxygen Therapy and Oxygen ToxicitydrsamnPas encore d'évaluation
- Arterial Blood Gases and Acid Base BalanceDocument12 pagesArterial Blood Gases and Acid Base BalancedrsamnPas encore d'évaluation
- Common Antibiotic Questions and TopicsDocument2 pagesCommon Antibiotic Questions and TopicsdrsamnPas encore d'évaluation
- Clinical Trials, 6 Principles: N.B. Patients Must Now Give Written Consent Before Entering A Clinical TrialDocument2 pagesClinical Trials, 6 Principles: N.B. Patients Must Now Give Written Consent Before Entering A Clinical TrialdrsamnPas encore d'évaluation
- Airway, Drips and ECGDocument17 pagesAirway, Drips and ECGdrsamnPas encore d'évaluation
- Science 2009-03-13Document112 pagesScience 2009-03-13drsamnPas encore d'évaluation
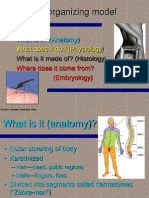
- Skin As An Organizing Model: What Is It? (Anatomy)Document11 pagesSkin As An Organizing Model: What Is It? (Anatomy)Chemist Mary MouresPas encore d'évaluation
- Layers of Abdominal Wall When Performing An AppendicectomyDocument2 pagesLayers of Abdominal Wall When Performing An AppendicectomydrsamnPas encore d'évaluation
- Lab FaqsDocument384 pagesLab FaqsPriyanka SinghPas encore d'évaluation
- Science 2009-01-30Document96 pagesScience 2009-01-30drsamnPas encore d'évaluation
- Impact Factor 2008Document119 pagesImpact Factor 2008abood127Pas encore d'évaluation
- Biology Under Construction - in Vitro Reconstitution of Cellular FunctionDocument8 pagesBiology Under Construction - in Vitro Reconstitution of Cellular FunctiondrsamnPas encore d'évaluation
- Midterm2 KeyDocument6 pagesMidterm2 KeyHungDoPas encore d'évaluation
- Predictive Ability of Social Cognitive Theory in Exercise Research: An Integrated Literature ReviewDocument13 pagesPredictive Ability of Social Cognitive Theory in Exercise Research: An Integrated Literature ReviewJovenlou BihagPas encore d'évaluation
- Catalog en Eurocoustic 0Document132 pagesCatalog en Eurocoustic 0Vikash KumarPas encore d'évaluation
- Numerical Modelling of Brine Dispersion in Shallow Coastal WatersDocument13 pagesNumerical Modelling of Brine Dispersion in Shallow Coastal WatersIAEME PublicationPas encore d'évaluation
- Sensus 143-80 Service Regulator R-1301 R5Document11 pagesSensus 143-80 Service Regulator R-1301 R5Paolita Auza BalderramaPas encore d'évaluation
- DBM MonetizationDocument2 pagesDBM MonetizationrsdiamzPas encore d'évaluation
- Dialysis and ElectrodialysisDocument32 pagesDialysis and ElectrodialysisJuan CarvajalPas encore d'évaluation
- Stanford - Trauma - Guidelines June 2016 Draft Adult and Peds FINALDocument166 pagesStanford - Trauma - Guidelines June 2016 Draft Adult and Peds FINALHaidir Muhammad100% (1)
- Fitting in and Fighting Back: Stigma Management Strategies Among Homeless KidsDocument24 pagesFitting in and Fighting Back: Stigma Management Strategies Among Homeless KidsIrisha AnandPas encore d'évaluation
- Rotating Driver S Workstation and Rotating Cab: Reliable Solutions For More Ergonomic and Effective WorkingDocument12 pagesRotating Driver S Workstation and Rotating Cab: Reliable Solutions For More Ergonomic and Effective WorkingEduardo CapeletiPas encore d'évaluation
- MAUS Catalogue PDFDocument10 pagesMAUS Catalogue PDFCarolina Garcés MoralesPas encore d'évaluation
- Guest-302344325-Quantitative Process AnalysisDocument4 pagesGuest-302344325-Quantitative Process AnalysisAhmedPas encore d'évaluation
- Preparation of Stick Type Solid Glue As Paper AdheDocument9 pagesPreparation of Stick Type Solid Glue As Paper AdheAhmad AlShahrourPas encore d'évaluation
- History of Crime and Criminal Justice in America EditedDocument7 pagesHistory of Crime and Criminal Justice in America EditedcarolinePas encore d'évaluation
- User Manual 4372070Document80 pagesUser Manual 4372070EDENILSON CORDEIROPas encore d'évaluation
- Nutrition Label ResponseDocument1 pageNutrition Label ResponseZoe JonesPas encore d'évaluation
- 2013 Medigate Profile PDFDocument26 pages2013 Medigate Profile PDFGabriel Duran DiazPas encore d'évaluation
- Fulltext PDFDocument55 pagesFulltext PDFManikandan VpPas encore d'évaluation
- Presented By: Agamjeet Singh Batra, VI - NDocument27 pagesPresented By: Agamjeet Singh Batra, VI - NlovleshrubyPas encore d'évaluation
- Equipment Available From SJ Turbine Inc.: LM Package Parts LM Package Parts High VoltageDocument59 pagesEquipment Available From SJ Turbine Inc.: LM Package Parts LM Package Parts High VoltageAnibal QuezadaPas encore d'évaluation
- Scala: Service InstructionsDocument16 pagesScala: Service Instructionsmario_turbinadoPas encore d'évaluation