Vous aimerez peut-être aussi
- Diagrama Somatizaciones - para Terapia IntegralDocument1 pageDiagrama Somatizaciones - para Terapia IntegralPERLA AGUILAR0% (1)
- Reflexologia Salud Con Masajes en Los Pies Lucia Roldan de Ortega 1Document65 pagesReflexologia Salud Con Masajes en Los Pies Lucia Roldan de Ortega 1Katya Flores Aduviri100% (1)
- Reap - RAZEDocument193 pagesReap - RAZECesar Antonio Tuanama Caro100% (1)
- Introducción A La Gastroenterología PDFDocument127 pagesIntroducción A La Gastroenterología PDFAlix pierina Chavez minoPas encore d'évaluation
- Principios Integrales Zoología PY098000Document42 pagesPrincipios Integrales Zoología PY098000Luis Humberto MZ67% (3)
- Cuadro Rocio!Document2 pagesCuadro Rocio!Brandon Lobos0% (1)
- La Hipertrofia Muscular y Su Entrenamiento Científico Más AvanzadoDocument53 pagesLa Hipertrofia Muscular y Su Entrenamiento Científico Más AvanzadoAlexander Asis González Olivares100% (2)
- MENINGOENCEFALITISDocument68 pagesMENINGOENCEFALITISRussell Hernan Huacho SusanivarPas encore d'évaluation
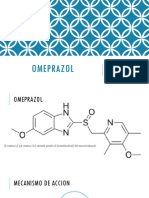
- Omeprazol ExposicionDocument10 pagesOmeprazol Exposicion徳利雅Pas encore d'évaluation
- Liceo Profesor Ladislao LeivaDocument50 pagesLiceo Profesor Ladislao LeivaBrandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiv11Document13 pagesLiceo Profesor Ladislao Leiv11Brandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiv11Document13 pagesLiceo Profesor Ladislao Leiv11Brandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiva MiaDocument27 pagesLiceo Profesor Ladislao Leiva MiaBrandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiv2Document54 pagesLiceo Profesor Ladislao Leiv2Brandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiv11Document13 pagesLiceo Profesor Ladislao Leiv11Brandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao LeivaDocument34 pagesLiceo Profesor Ladislao LeivaBrandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiva equipo de trabajo tecnologíaDocument29 pagesLiceo Profesor Ladislao Leiva equipo de trabajo tecnologíaBrandon LobosPas encore d'évaluation
- Caratula Tarea 2Document3 pagesCaratula Tarea 2Brandon LobosPas encore d'évaluation
- Liceo Profesor Ladislao Leiva222Document28 pagesLiceo Profesor Ladislao Leiva222Brandon LobosPas encore d'évaluation
- Mi TareaDocument18 pagesMi TareaBrandon LobosPas encore d'évaluation
- Curriculum 1Document2 pagesCurriculum 1Brandon LobosPas encore d'évaluation
- Caratula Tarea 2Document3 pagesCaratula Tarea 2Brandon LobosPas encore d'évaluation
- Cuestionario de FilosofiaDocument1 pageCuestionario de FilosofiaBrandon LobosPas encore d'évaluation
- Especificidad de ProteinasDocument7 pagesEspecificidad de ProteinasBrandon LobosPas encore d'évaluation
- Caratula Tarea 2Document3 pagesCaratula Tarea 2Brandon LobosPas encore d'évaluation
- Redes Neuronales en RDocument31 pagesRedes Neuronales en RquispeluisPas encore d'évaluation
- Fibromialgia y Fisioterapia Global PDFDocument5 pagesFibromialgia y Fisioterapia Global PDFTephi Gomez ImaraiPas encore d'évaluation
- Tarea 2 Hilda JosefinaDocument4 pagesTarea 2 Hilda JosefinaTania GonzalezPas encore d'évaluation
- ESQUEMA Aparato Respiratorio FiniquitoDocument1 pageESQUEMA Aparato Respiratorio FiniquitoAnnaPas encore d'évaluation
- Comunicación No VerbalDocument25 pagesComunicación No VerbalBubulina Ramírez Llera100% (1)
- Trabajo Práctico REFLEJOS MEDULARES PDFDocument12 pagesTrabajo Práctico REFLEJOS MEDULARES PDFSabrina PortelliPas encore d'évaluation
- Conducto InguinalDocument8 pagesConducto InguinalJosbel Alejandro RodríguezPas encore d'évaluation
- A Mecanica CorporalDocument21 pagesA Mecanica CorporalAnel CastilloPas encore d'évaluation
- Preguntas Concurso de BiologiaDocument40 pagesPreguntas Concurso de BiologiaLESLIE PAULETTEPas encore d'évaluation
- Enfermedad Arterial Obstructiva CrónicaDocument21 pagesEnfermedad Arterial Obstructiva CrónicaMartina Alarcon AcevedoPas encore d'évaluation
- Pregunta 4 y 5 RenalDocument6 pagesPregunta 4 y 5 RenalchristianrojashPas encore d'évaluation
- BORRADOR - INFORME DE NECROPSIA - HUEMUL - G - AprileDocument10 pagesBORRADOR - INFORME DE NECROPSIA - HUEMUL - G - Aprilecorreopuertae8824Pas encore d'évaluation
- Nro Tal ID Altura Tipo Taladro Hoja Tipo EmulsionDocument14 pagesNro Tal ID Altura Tipo Taladro Hoja Tipo EmulsionJohn Jairo Cheppe AguirrePas encore d'évaluation
- Flagelos eucariotas: estructura y funciónDocument4 pagesFlagelos eucariotas: estructura y funciónRicardo SandovalPas encore d'évaluation
- DE 50 Sedante 1Document9 pagesDE 50 Sedante 1JulioCeesaarGPas encore d'évaluation
- Anguila Eléctrica: Pez que emite descargas de 600VDocument13 pagesAnguila Eléctrica: Pez que emite descargas de 600VPilar Oversluijs QuevedoPas encore d'évaluation
- Introducción Al Ciclo de Vida de Las AvesDocument95 pagesIntroducción Al Ciclo de Vida de Las AvesJesus J. Zuñiga100% (1)
- Formal I ZarDocument2 pagesFormal I ZarMARIOPas encore d'évaluation
- Los Linfocitos B y La Inmunidad Humoral PDFDocument13 pagesLos Linfocitos B y La Inmunidad Humoral PDFKarla Pam LópezPas encore d'évaluation
- Triptico KionDocument2 pagesTriptico KionalePas encore d'évaluation
- Hemograma automatizado: 18 parámetrosDocument49 pagesHemograma automatizado: 18 parámetrosYesenia Aide Vivar AlvaPas encore d'évaluation