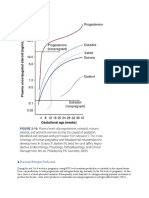
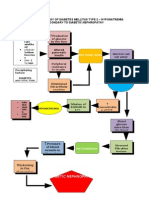
Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5782)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Melo, Moinca T. Case Reading Vaginal Bleeding Causes Patient Dysfunctional Uterine BleedingDocument2 pagesMelo, Moinca T. Case Reading Vaginal Bleeding Causes Patient Dysfunctional Uterine BleedingMonica Melo HernandezPas encore d'évaluation
- Healthcare of The Transgender Patient - Version 5.3 (Read-Only)Document117 pagesHealthcare of The Transgender Patient - Version 5.3 (Read-Only)Prasanna AyerPas encore d'évaluation
- Green Algae May Help Female FertilityDocument10 pagesGreen Algae May Help Female FertilityBulgar.Pas encore d'évaluation
- Gynecological NursingDocument329 pagesGynecological Nursingsharon ocharaPas encore d'évaluation
- Thyroidectomy: Continuing Education ActivityDocument10 pagesThyroidectomy: Continuing Education Activitybo gum parkPas encore d'évaluation
- Abnormal Uterine BleedingDocument36 pagesAbnormal Uterine BleedingPranshu Prajyot 67100% (1)
- Science Discovers The Physiological Value of Continence 2Document20 pagesScience Discovers The Physiological Value of Continence 2shaswata_panjaPas encore d'évaluation
- Adaptogen Herbs to Lower Stress Hormone CortisolDocument3 pagesAdaptogen Herbs to Lower Stress Hormone CortisolJames ConnerPas encore d'évaluation
- Estrogen DR DeanDocument8 pagesEstrogen DR DeanMochammad Halim NPas encore d'évaluation
- .Document544 pages.Bestiana SaraLiontina0% (1)
- Diabetes Questionnaire GuideDocument1 pageDiabetes Questionnaire Guideashmeet bansalPas encore d'évaluation
- ASRM - Diagnostic Evaluation of The Infertile FemaleDocument11 pagesASRM - Diagnostic Evaluation of The Infertile FemaleMina zhouPas encore d'évaluation
- TRAb MaglumiDocument6 pagesTRAb MaglumiLaboratoire Dr Mansouri Reghaia AlgerPas encore d'évaluation
- Anatomy and Physiology of Farm AnimalsDocument157 pagesAnatomy and Physiology of Farm AnimalsOliver TalipPas encore d'évaluation
- Chronic Kidney Disease-Mineral Bone Disease (CKD-MBD) : What Is Renal Bone Disease? How Is It Diagnosed?Document1 pageChronic Kidney Disease-Mineral Bone Disease (CKD-MBD) : What Is Renal Bone Disease? How Is It Diagnosed?Gabriel VargasPas encore d'évaluation
- DR Shahid Manzur: Associate Professor Department of Diagnostic Radiology BVHDocument24 pagesDR Shahid Manzur: Associate Professor Department of Diagnostic Radiology BVHdrqazi777Pas encore d'évaluation
- Pathophysiology of Diabetes Mellitus Type 2Document1 pagePathophysiology of Diabetes Mellitus Type 2faula rocamora100% (3)
- Calcium Homeostasis: Endocrine Regulation of (Ca)Document4 pagesCalcium Homeostasis: Endocrine Regulation of (Ca)PRANAB KUMAR MUKHERJEEPas encore d'évaluation
- DQC20 Depliant 4vol DiabeteDeGrossesse EN WebDocument2 pagesDQC20 Depliant 4vol DiabeteDeGrossesse EN WebZH. omg sarPas encore d'évaluation
- Anatomy & Physiology: Male Reproductive SystemDocument7 pagesAnatomy & Physiology: Male Reproductive Systemkristel ludangcoPas encore d'évaluation
- Hypertension: Basic InformationDocument8 pagesHypertension: Basic InformationHector OrtegaPas encore d'évaluation
- 2 Glucose PracticalDocument20 pages2 Glucose PracticalMada SaadPas encore d'évaluation
- Arterial Blood Pressure RegulationDocument21 pagesArterial Blood Pressure Regulationnasihtas100% (2)
- CarbimazoleDocument3 pagesCarbimazoleManish SharmaPas encore d'évaluation
- Case Study: Carol: By: Manpreet Kaur, Sarah Riley, Ben AvilaDocument14 pagesCase Study: Carol: By: Manpreet Kaur, Sarah Riley, Ben AvilaBuchet AmaliePas encore d'évaluation
- Ovulation Test: How Clearblue® DIGITAL Ovulation Test Works With YouDocument2 pagesOvulation Test: How Clearblue® DIGITAL Ovulation Test Works With YoutadiganeshPas encore d'évaluation
- Syndromes of Ketosis-Prone Diabetes Mellitus - UpToDateDocument17 pagesSyndromes of Ketosis-Prone Diabetes Mellitus - UpToDateDanPas encore d'évaluation
- Hypogonadotropic Hypogonadism and Gynaecomastia in The Young Adult: A Case SeriesDocument4 pagesHypogonadotropic Hypogonadism and Gynaecomastia in The Young Adult: A Case Seriesricepaddy1867Pas encore d'évaluation
- DLC911 - Feedback Mechanism Lesson PlanDocument2 pagesDLC911 - Feedback Mechanism Lesson PlanMariel CondesaPas encore d'évaluation
- Renal System Physiology: GFR, Tubular Reabsorption and Urine ConcentrationDocument47 pagesRenal System Physiology: GFR, Tubular Reabsorption and Urine ConcentrationMuhammad Saim Bin Saeed0% (1)