Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Drug ListDocument127 pagesDrug Listnehal choudharyPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Astm B733-04 (2014)Document5 pagesAstm B733-04 (2014)vtsusr fvPas encore d'évaluation
- FAG-Jaw Crusher BearingsDocument38 pagesFAG-Jaw Crusher BearingsJinto A J100% (2)
- ASME PTC 19.5-2004 TocDocument9 pagesASME PTC 19.5-2004 TocVamsi Yashoda0% (2)
- 5 PlasterDocument3 pages5 PlasterMuhammad AgungPas encore d'évaluation
- Complexmetric TitrationDocument17 pagesComplexmetric TitrationAnonymous oC3F7cxlLH100% (1)
- L&T Ball ValveDocument12 pagesL&T Ball ValvecdsenthilPas encore d'évaluation
- Hammer CrusherDocument4 pagesHammer CrusherkhoaglePas encore d'évaluation
- Comparison Between Currently Applied Grinding TechnologiesDocument28 pagesComparison Between Currently Applied Grinding TechnologiesSai Sricharan Reddy100% (3)
- PREHEATER OptimizationDocument2 pagesPREHEATER OptimizationsaeedhoseiniPas encore d'évaluation
- Astm D129 PDFDocument4 pagesAstm D129 PDFProkop100% (1)
- Modular Air Cooled Chillers PDFDocument22 pagesModular Air Cooled Chillers PDFBehnamPas encore d'évaluation
- Optimisation of The Rotary KilnDocument34 pagesOptimisation of The Rotary KilnSampathkumar Mtech100% (3)
- Handbook On Chlorine HandlingDocument106 pagesHandbook On Chlorine Handlingkirandevi1981100% (1)
- Generating Surface Crack PatternsDocument11 pagesGenerating Surface Crack PatternskhoaglePas encore d'évaluation
- Homework 2 - SolutionDocument3 pagesHomework 2 - SolutionkhoaglePas encore d'évaluation
- Useful PhrasesDocument8 pagesUseful PhraseswinnieXDPas encore d'évaluation
- Causes of Crackle On Porcelain WareDocument2 pagesCauses of Crackle On Porcelain WarekhoaglePas encore d'évaluation
- Journal Paper IJAETDocument8 pagesJournal Paper IJAETvempada1942Pas encore d'évaluation
- Lecture 2Document4 pagesLecture 2khoaglePas encore d'évaluation
- Choosing The Right Motors For Your Mills: Alex Doll and Derek BarrattDocument16 pagesChoosing The Right Motors For Your Mills: Alex Doll and Derek Barrattavca65Pas encore d'évaluation
- Airflex Grinding MillsDocument6 pagesAirflex Grinding MillskhoaglePas encore d'évaluation
- CA501Document614 pagesCA501Ignacio AntonioPas encore d'évaluation
- Electric Mortor For Ball MillDocument2 pagesElectric Mortor For Ball MillkhoaglePas encore d'évaluation
- United States Patent (191: Andersen Et A1Document5 pagesUnited States Patent (191: Andersen Et A1khoaglePas encore d'évaluation
- United States Patent (191: Andersen Et A1Document5 pagesUnited States Patent (191: Andersen Et A1khoaglePas encore d'évaluation
- Effect of Curing Temperature On The Development of Hard Structure of Metakaolin-Based GeopolymerDocument8 pagesEffect of Curing Temperature On The Development of Hard Structure of Metakaolin-Based GeopolymerkhoaglePas encore d'évaluation
- Phu Luc 3Document2 pagesPhu Luc 3khoaglePas encore d'évaluation
- Properties of Metakaolin Geopolymer Hardened Paste Prepared by High-Pressure CompactionDocument8 pagesProperties of Metakaolin Geopolymer Hardened Paste Prepared by High-Pressure CompactionkhoaglePas encore d'évaluation
- Effect of Porosity On The Absorbed, Reemitted and Transmitted Light by A Geopolymer Metakaolin BaseDocument4 pagesEffect of Porosity On The Absorbed, Reemitted and Transmitted Light by A Geopolymer Metakaolin BasekhoaglePas encore d'évaluation
- Electrostatic Precipitator - Mechanical DescriptionDocument19 pagesElectrostatic Precipitator - Mechanical Descriptionhozipek5599Pas encore d'évaluation
- 1Document6 pages1basanth_712982Pas encore d'évaluation
- PaulingDocument4 pagesPaulingkhoaglePas encore d'évaluation
- NDT NLU Detection Micro Cracks ConcreteDocument7 pagesNDT NLU Detection Micro Cracks ConcreteElango PaulchamyPas encore d'évaluation
- ConvertDocument8 pagesConvertkhoaglePas encore d'évaluation
- Pauling RulesDocument5 pagesPauling RuleskhoaglePas encore d'évaluation
- Calcium Carbonate Precipitation by Different Bacterial StrainsDocument14 pagesCalcium Carbonate Precipitation by Different Bacterial StrainskhoaglePas encore d'évaluation
- Unit 1Document29 pagesUnit 1Vasanth AradhyaPas encore d'évaluation
- BindersDocument4 pagesBindersSariyyaHeydarovaPas encore d'évaluation
- Beetroot IA JilleanDocument10 pagesBeetroot IA Jilleanzeen2011Pas encore d'évaluation
- HepasilDocument1 pageHepasilDea Angela BelmontePas encore d'évaluation
- SirCar 2002Document4 pagesSirCar 2002Zahra GhPas encore d'évaluation
- Dyeing Fault .Document22 pagesDyeing Fault .alimamit43Pas encore d'évaluation
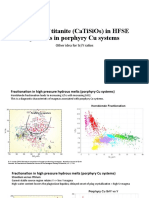
- The Role of Titanite (CaTiSiO ) in PCD ExplorationDocument9 pagesThe Role of Titanite (CaTiSiO ) in PCD ExplorationMarco Augusto Robles AncajimaPas encore d'évaluation
- Composicion Agar NNNDocument2 pagesComposicion Agar NNNjhrpaul15Pas encore d'évaluation
- HistotechDocument46 pagesHistotechviolaorsino100% (1)
- DP1SL Redox UT VCDocument6 pagesDP1SL Redox UT VCmarilee huntPas encore d'évaluation
- M1226e Peristaltic Pump Compact b19b PDFDocument12 pagesM1226e Peristaltic Pump Compact b19b PDFRenatoPas encore d'évaluation
- Tube Erosion PDFDocument9 pagesTube Erosion PDFktjayakumar3878Pas encore d'évaluation
- Difference Between Paper and Vs Thin Layer and Vs Column ChromatographyDocument8 pagesDifference Between Paper and Vs Thin Layer and Vs Column ChromatographySubhradeep GhoshPas encore d'évaluation
- Reactions and Stoichiometry Cheat Sheet: by ViaDocument3 pagesReactions and Stoichiometry Cheat Sheet: by ViaNeia De JesusPas encore d'évaluation
- Effect of Eggshell and Silk Fibroin On StyreneDocument1 pageEffect of Eggshell and Silk Fibroin On Styreneangeleekaye abelindePas encore d'évaluation
- Square Planar Substitution and Trans Effect-2Document10 pagesSquare Planar Substitution and Trans Effect-2aliyyaPas encore d'évaluation
- Nano-Particle Prepared From Sol-Gel Method: Titanium Tetra-Iso-Propoxide DiethanolamineDocument21 pagesNano-Particle Prepared From Sol-Gel Method: Titanium Tetra-Iso-Propoxide DiethanolamineMaggyBalcazarPas encore d'évaluation
- Dow Membrane CleaningDocument2 pagesDow Membrane CleaningDonatas Bertasius100% (1)
- Difcomanualofdeh 09 DigeDocument360 pagesDifcomanualofdeh 09 DigeDimas FirliantoroPas encore d'évaluation
- Mil PRF 8565kDocument39 pagesMil PRF 8565kgustavojorge12Pas encore d'évaluation
- Material Safety Data Sheet: I - Product IdentificationDocument2 pagesMaterial Safety Data Sheet: I - Product IdentificationAndini Nur PaujiahPas encore d'évaluation