Vous aimerez peut-être aussi
- Biogeo11 Teste4Document9 pagesBiogeo11 Teste4crisvit90% (10)
- Catálogo Neodent 2015Document210 pagesCatálogo Neodent 2015Larissa Sestari100% (1)
- Apostila Bioquimica 2Document112 pagesApostila Bioquimica 2_nei100% (6)
- Planilha Treino 21k - Iniciantes-NvDocument2 pagesPlanilha Treino 21k - Iniciantes-NvGustavo Bratfisch100% (7)
- Exercícios de Revisão Com Respostas - GEODocument12 pagesExercícios de Revisão Com Respostas - GEOSilvestre Gonçalves de Souza100% (1)
- A História Do Cristianismo e de Suas EscriturasDocument28 pagesA História Do Cristianismo e de Suas EscriturasAdv AlmeidaPas encore d'évaluation
- QuilotóraxDocument7 pagesQuilotóraxMárcia NPPas encore d'évaluation
- IGREJA Construtora Renovadora Formadora Da CivilizaçãoDocument15 pagesIGREJA Construtora Renovadora Formadora Da CivilizaçãoRaul MarcelPas encore d'évaluation
- Pages From RN 09 02-4Document10 pagesPages From RN 09 02-4Silvestre Gonçalves de SouzaPas encore d'évaluation
- Manual ExamesDocument82 pagesManual ExamesSilvestre Gonçalves de Souza67% (3)
- Derrames Pleurais Fisiopatologia DiagnosticoDocument8 pagesDerrames Pleurais Fisiopatologia DiagnosticoSilvestre Gonçalves de SouzaPas encore d'évaluation
- ParasitologiaDocument49 pagesParasitologiaAleCarvalho1100% (1)
- IMUNOHEMATOLOGIADocument3 pagesIMUNOHEMATOLOGIAWeriston Alves100% (1)
- Dna RnaDocument8 pagesDna RnaJainny FerreiraPas encore d'évaluation
- As Idéias de EinsteinDocument142 pagesAs Idéias de EinsteinFelipe00960% (1)
- Aprendizagem - Teorias e ProcessosDocument107 pagesAprendizagem - Teorias e ProcessosNeuropsico pedagogiaPas encore d'évaluation
- Manual Do Aluno IPMIL Versao 2Document52 pagesManual Do Aluno IPMIL Versao 2Manuela EspinosaPas encore d'évaluation
- Referencial de Formação Tecnológica - TECDocument5 pagesReferencial de Formação Tecnológica - TECadelinafelizardoPas encore d'évaluation
- Alexandre Ordem Paranormal Wiki FandomDocument2 pagesAlexandre Ordem Paranormal Wiki FandomIevan polkaPas encore d'évaluation
- Revisão 9ano 3triDocument4 pagesRevisão 9ano 3trieduarda.cavalcantigsPas encore d'évaluation
- História Dos Brinquedos - BarbieDocument2 pagesHistória Dos Brinquedos - BarbieMarcielly TavaresPas encore d'évaluation
- Cap. 04 - Geometria, Ideias Intuitivas.Document3 pagesCap. 04 - Geometria, Ideias Intuitivas.Douglas OliveiraPas encore d'évaluation
- Educacao Infantil Como Organizar o Espaco Escolar Com Intencionalidade PedagogicaDocument6 pagesEducacao Infantil Como Organizar o Espaco Escolar Com Intencionalidade PedagogicaIsa OliveiraPas encore d'évaluation
- Dislexia - Vol 3Document32 pagesDislexia - Vol 3Ariel MedeirosPas encore d'évaluation
- Socorrista - AphDocument67 pagesSocorrista - AphJonas AquinoPas encore d'évaluation
- Bolsas e Sapatos (20 Artigos)Document30 pagesBolsas e Sapatos (20 Artigos)Maira SiqueiraPas encore d'évaluation
- 2 Regras Do ArDocument26 pages2 Regras Do ArJota JuniorPas encore d'évaluation
- Hipnose Na Prática - Um Guia Com Tudo Que Você Precisa SaberDocument15 pagesHipnose Na Prática - Um Guia Com Tudo Que Você Precisa SaberCleide Helena MacedoPas encore d'évaluation
- Vitrine Tupperware 1.2017Document68 pagesVitrine Tupperware 1.2017Tupperware ShowPas encore d'évaluation
- S - VelocidadesDocument5 pagesS - Velocidadesjoao piter de paulaPas encore d'évaluation
- Modulo2 - Modulo de Resistencia A FlexaoDocument13 pagesModulo2 - Modulo de Resistencia A FlexaoJéssika PauloPas encore d'évaluation
- A Epistemologia e A Psicologia Genética de Jean PiagetDocument22 pagesA Epistemologia e A Psicologia Genética de Jean PiagetHiago Carvalho100% (1)
- SEMANÁRIO 5 º ANO 23 A 27 de FevereiroDocument12 pagesSEMANÁRIO 5 º ANO 23 A 27 de FevereiroClaudia LeitePas encore d'évaluation
- 1a Lista de ExercíciosDocument2 pages1a Lista de ExercíciosEvandro CKPas encore d'évaluation
- O Abuso Sexual, Estudo de Casos em Cenas Incestuosas PDFDocument9 pagesO Abuso Sexual, Estudo de Casos em Cenas Incestuosas PDFAléxia VegaPas encore d'évaluation
- Manual de ProjetosDocument226 pagesManual de ProjetosbetobragajrPas encore d'évaluation
- I CZC - Livro Do Evento - 2016-With-Cover-Page-V2Document232 pagesI CZC - Livro Do Evento - 2016-With-Cover-Page-V2Joao FelipePas encore d'évaluation
- Aso Carlos GustavoDocument1 pageAso Carlos GustavowescleiPas encore d'évaluation
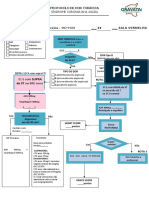
- Protocolo de Dor TorácicaDocument2 pagesProtocolo de Dor Torácicamedicos.upamoradasPas encore d'évaluation
- Mapa Mental Tecido MuscularDocument1 pageMapa Mental Tecido MuscularandressalopesmmPas encore d'évaluation
- Geografia - Diversidade ÉtnicaDocument7 pagesGeografia - Diversidade ÉtnicaJosue felipe BarbosaPas encore d'évaluation