Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Concise Human Body Book - New Edition by DKDocument322 pagesThe Concise Human Body Book - New Edition by DKJim RenniePas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
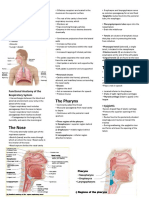
- The Respiratory System ReviewerDocument6 pagesThe Respiratory System ReviewerJP LozadaPas encore d'évaluation
- Materi 4 - Kanker Pada Perempuan - DR - Tofan, SP - Og (K)Document41 pagesMateri 4 - Kanker Pada Perempuan - DR - Tofan, SP - Og (K)Tuti KristantiPas encore d'évaluation
- Disorders of The Thyroid GlandDocument21 pagesDisorders of The Thyroid GlandAgustina TambingPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Urinary SystemDocument9 pagesUrinary SystemCailah Sofia SelausoPas encore d'évaluation
- IABP Learning PackageDocument35 pagesIABP Learning PackageMuhammad Mahbøøb SadiqPas encore d'évaluation
- Urinary System Nuclear Medicine: Renal ImagingDocument68 pagesUrinary System Nuclear Medicine: Renal Imagingapi-19916399Pas encore d'évaluation
- Chapter 1 NotesDocument5 pagesChapter 1 NotesKatarina GruborovićPas encore d'évaluation
- Mammary Gland Development - Methods and Protocols PDFDocument357 pagesMammary Gland Development - Methods and Protocols PDFMihaela FloreaPas encore d'évaluation
- 7 - The Biliary TractDocument48 pages7 - The Biliary TractKim RamosPas encore d'évaluation
- Psyc1000 1.1 Textbook NotesDocument3 pagesPsyc1000 1.1 Textbook NotesKatarina GruborovićPas encore d'évaluation
- PSYC 2410 de S12 Textbook Notes Chapter 02Document15 pagesPSYC 2410 de S12 Textbook Notes Chapter 02Katarina GruborovićPas encore d'évaluation
- How Drugs Impact the Brain's Reward SystemDocument22 pagesHow Drugs Impact the Brain's Reward SystemKatarina GruborovićPas encore d'évaluation
- PSYC 2410 DE S12 Textbook Notes Chapter 08 PDFDocument12 pagesPSYC 2410 DE S12 Textbook Notes Chapter 08 PDFKatarina GruborovićPas encore d'évaluation
- PSYC 2410 DE S12 Textbook Notes Chapter 07 PDFDocument12 pagesPSYC 2410 DE S12 Textbook Notes Chapter 07 PDFKatarina GruborovićPas encore d'évaluation
- PSYC 2410 DE S12 Textbook Notes Chapter 04 PDFDocument14 pagesPSYC 2410 DE S12 Textbook Notes Chapter 04 PDFKatarina GruborovićPas encore d'évaluation
- PSYC 2410 DE S12 Textbook Notes Chapter 05 PDFDocument17 pagesPSYC 2410 DE S12 Textbook Notes Chapter 05 PDFKatarina GruborovićPas encore d'évaluation
- PSYC 2410 de S12 Textbook Notes Chapter 02Document15 pagesPSYC 2410 de S12 Textbook Notes Chapter 02Katarina GruborovićPas encore d'évaluation
- PSYC 2410 DE S12 Textbook Notes Chapter 03 PDFDocument11 pagesPSYC 2410 DE S12 Textbook Notes Chapter 03 PDFKatarina GruborovićPas encore d'évaluation
- Factors Affecting Renal ExcretionDocument19 pagesFactors Affecting Renal ExcretionV. SravaniPas encore d'évaluation
- Case WriteupDocument5 pagesCase WriteupBatool AldaherPas encore d'évaluation
- Chapter 6 - IntegumentaryDocument63 pagesChapter 6 - Integumentaryapi-297069611Pas encore d'évaluation
- Gatehouse Report PDFDocument978 pagesGatehouse Report PDFTauseek RangrejPas encore d'évaluation
- A Gastro Tahun Lalu - Tahun IniDocument12 pagesA Gastro Tahun Lalu - Tahun IniDapot SianiparPas encore d'évaluation
- Mabini Colleges Provides Quality EducationDocument2 pagesMabini Colleges Provides Quality Educationkenneth loPas encore d'évaluation
- ThyroidectomyDocument2 pagesThyroidectomykzone2290Pas encore d'évaluation
- DermatopatologiDocument117 pagesDermatopatologiSariPas encore d'évaluation
- Hormones & GlandsDocument3 pagesHormones & GlandsMickaela Delos SantosPas encore d'évaluation
- Disease: Community Acquired PneumoniaDocument4 pagesDisease: Community Acquired PneumoniaJo Traven AzueloPas encore d'évaluation
- The Clinical Significance of Subclinical Thyroid Dysfunction Bernadette Biondi and David S. Cooper PDFDocument56 pagesThe Clinical Significance of Subclinical Thyroid Dysfunction Bernadette Biondi and David S. Cooper PDFLorena IbarrolaPas encore d'évaluation
- Anatomical and Physiological ChangesDocument25 pagesAnatomical and Physiological ChangespoojasolPas encore d'évaluation
- SurgeryDocument146 pagesSurgeryNaren ShanPas encore d'évaluation
- Digestive and Gastrointestinal Function HandoutsDocument15 pagesDigestive and Gastrointestinal Function HandoutsChristine Joy FloresPas encore d'évaluation
- Tortora 13e Glossary 2012Document30 pagesTortora 13e Glossary 2012Shweta KhannaPas encore d'évaluation
- Endocrine NotesDocument9 pagesEndocrine NotesLucky GomezPas encore d'évaluation
- Nsci Exam 3 Study SheetDocument8 pagesNsci Exam 3 Study SheetHan TaubPas encore d'évaluation
- Jurnal 4 Uji Jalan 6 MenitDocument10 pagesJurnal 4 Uji Jalan 6 MenitSandy Tama Okvan YudhaPas encore d'évaluation
- Circulatory SystemDocument20 pagesCirculatory SystemsuryanshPas encore d'évaluation
- Reading As A Neuro-Physiological ProcessDocument17 pagesReading As A Neuro-Physiological ProcessStephanie BuenafePas encore d'évaluation
- Mills 1975Document6 pagesMills 1975Nida AtmimPas encore d'évaluation