Vous aimerez peut-être aussi
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Promotions & Design 2009Document153 pagesPromotions & Design 2009Lakis Takis PapasPas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Metro Ultra FreshDocument2 pagesMetro Ultra FreshLakis Takis PapasPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Metro Gold 1215Document16 pagesMetro Gold 1215Lakis Takis PapasPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Metro Traders 1215Document10 pagesMetro Traders 1215Lakis Takis PapasPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- OUM 29 PenDocument1 pageOUM 29 PenLakis Takis PapasPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Top 20 Cancer DrugsDocument8 pagesTop 20 Cancer DrugsLakis Takis PapasPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Metro Traders 1215Document10 pagesMetro Traders 1215Lakis Takis PapasPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Metro Food 1215Document31 pagesMetro Food 1215Lakis Takis PapasPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- TMRDocument11 pagesTMRTanisha FlowersPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Junyu Industrial Group: Fresh Milk DispenserDocument6 pagesJunyu Industrial Group: Fresh Milk DispenserLakis Takis PapasPas encore d'évaluation
- Metro Aro 1215Document17 pagesMetro Aro 1215Lakis Takis PapasPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Tardo Filipinas, Inc. Company ProfileDocument3 pagesTardo Filipinas, Inc. Company ProfileLakis Takis PapasPas encore d'évaluation
- Metro 5 Crazy Days 1215Document12 pagesMetro 5 Crazy Days 1215Lakis Takis PapasPas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Chison Q5 BrochureDocument8 pagesChison Q5 BrochureLakis Takis PapasPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Catalogue Product NewDocument6 pagesCatalogue Product NewLakis Takis PapasPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- ΒΑΡΟΥΦΑΚΗς PDFDocument8 pagesΒΑΡΟΥΦΑΚΗς PDFLakis Takis PapasPas encore d'évaluation
- Russian VodkaDocument13 pagesRussian VodkaLakis Takis PapasPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Confectionery and ChocolateDocument18 pagesConfectionery and ChocolateLakis Takis PapasPas encore d'évaluation
- Companydetails Logistic Partner Parent and Subsidariy CompanyDocument1 pageCompanydetails Logistic Partner Parent and Subsidariy CompanyLakis Takis PapasPas encore d'évaluation
- Altair Trade LTD.: 5, Office, 70, Dolomanovskiy Alley, Rostov-On-Don, Russia, 344011Document16 pagesAltair Trade LTD.: 5, Office, 70, Dolomanovskiy Alley, Rostov-On-Don, Russia, 344011Lakis Takis PapasPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Wine From KUBANDocument10 pagesWine From KUBANLakis Takis PapasPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Pridvin e Product CatalogueDocument26 pagesPridvin e Product CatalogueLakis Takis PapasPas encore d'évaluation
- Wine From Kuban1Document6 pagesWine From Kuban1Lakis Takis PapasPas encore d'évaluation
- Companydetails Logistic Partner Parent and Subsidariy CompanyDocument1 pageCompanydetails Logistic Partner Parent and Subsidariy CompanyLakis Takis PapasPas encore d'évaluation
- Companydetails Logistic Partner Parent and Subsidariy CompanyDocument1 pageCompanydetails Logistic Partner Parent and Subsidariy CompanyLakis Takis PapasPas encore d'évaluation
- Dauco 2014 ING - BL PDFDocument1 pageDauco 2014 ING - BL PDFLakis Takis PapasPas encore d'évaluation
- Keg Lager Cider ListDocument3 pagesKeg Lager Cider ListLakis Takis PapasPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Ron BaraimaDocument4 pagesRon BaraimaLakis Takis PapasPas encore d'évaluation
- Tds Thracia 2010 enDocument1 pageTds Thracia 2010 enLakis Takis PapasPas encore d'évaluation
- 13 ChromosomesDocument36 pages13 ChromosomesKiki QqsPas encore d'évaluation
- Biotechnology Unzipped Promises and Realities - Eric S. GraceDocument264 pagesBiotechnology Unzipped Promises and Realities - Eric S. GraceMayank Pathak100% (1)
- Maximum Dosage PolicyDocument10 pagesMaximum Dosage PolicyTosy SatyoPas encore d'évaluation
- A Collagen Peptide-Based Physical Hydrogel ForDocument6 pagesA Collagen Peptide-Based Physical Hydrogel Forsuri_2113Pas encore d'évaluation
- Regional Director Biotech Sales in Minneapolis ST Paul Metro MN Resume Mark BeenkenDocument2 pagesRegional Director Biotech Sales in Minneapolis ST Paul Metro MN Resume Mark BeenkenMark BeenkenPas encore d'évaluation
- Prokaryote Vs EukaryoteDocument19 pagesProkaryote Vs Eukaryoteapi-302494239Pas encore d'évaluation
- ARID1A Mutations in Endometriosis-Associated Ovarian: CarcinomasDocument6 pagesARID1A Mutations in Endometriosis-Associated Ovarian: CarcinomasherryPas encore d'évaluation
- A Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate IndependentlyDocument5 pagesA Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate Independentlyyaqoob008Pas encore d'évaluation
- Snep PDFDocument13 pagesSnep PDFNaga RajanPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Genscript Annual Report 2018Document73 pagesGenscript Annual Report 2018oaifoiweuPas encore d'évaluation
- Final KeyDocument11 pagesFinal KeyAbhinandan ChakrabortyPas encore d'évaluation
- A Study On Protein Sequence Clustering With OPTICSDocument6 pagesA Study On Protein Sequence Clustering With OPTICSJournal of ComputingPas encore d'évaluation
- Definitive PCRDocument55 pagesDefinitive PCRSumit MitraPas encore d'évaluation
- Biotechnology Sample Sop 3Document3 pagesBiotechnology Sample Sop 3api-3816734100% (2)
- CV - Dr. A.G. Ingale PDFDocument8 pagesCV - Dr. A.G. Ingale PDFIntegrated Intelligent ResearchPas encore d'évaluation
- Grand Challenges India Application InstructionsDocument2 pagesGrand Challenges India Application InstructionsSulekha BhattacherjeePas encore d'évaluation
- Mic MBCDocument5 pagesMic MBCOneil CerdenaPas encore d'évaluation
- Total Synthesis of Escherichia Coli With A Recoded GenomeDocument20 pagesTotal Synthesis of Escherichia Coli With A Recoded GenomeEstalin CaizapantaPas encore d'évaluation
- Gram-Negative Bacteria - Wikipedia, The Free EncyclopediaDocument8 pagesGram-Negative Bacteria - Wikipedia, The Free EncyclopediaNadiya AfifahPas encore d'évaluation
- I 10 GandjarDocument6 pagesI 10 GandjarDio AugiePas encore d'évaluation
- Beginning Molecular Biology Laboratory Manual-UMBC.2003 (Por Trisquelion)Document52 pagesBeginning Molecular Biology Laboratory Manual-UMBC.2003 (Por Trisquelion)Sivan AndiPas encore d'évaluation
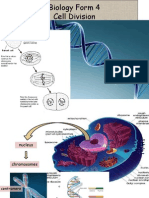
- Biology Form 4Document13 pagesBiology Form 4Arvinth Guna SegaranPas encore d'évaluation
- Journals: Department of Chemical EngineeringDocument26 pagesJournals: Department of Chemical EngineeringshankyforuPas encore d'évaluation
- Anti-Adhesion and Antibiotic Modulatory Evaluation of Grapefruit (Citrus Paradisi) Juice and Seed Extract On Bacteria Isolated From Urine and CatheterDocument14 pagesAnti-Adhesion and Antibiotic Modulatory Evaluation of Grapefruit (Citrus Paradisi) Juice and Seed Extract On Bacteria Isolated From Urine and CatheterJournal of Pharmacy & Pharmacognosy ResearchPas encore d'évaluation
- Kishore AccountsDocument4 pagesKishore AccountskishorePas encore d'évaluation
- Cellobiose Dehydrogenase and A Copper-Dependent Polysaccharide Monooxigenase Potentiate Cellulose Degradation by Neurospora CrassaDocument8 pagesCellobiose Dehydrogenase and A Copper-Dependent Polysaccharide Monooxigenase Potentiate Cellulose Degradation by Neurospora CrassaRoberto MaedaPas encore d'évaluation
- Essay - DnaDocument2 pagesEssay - Dnaapi-243852896Pas encore d'évaluation
- Outlook-2020 PDFDocument116 pagesOutlook-2020 PDFmatlabbPas encore d'évaluation
- Mode of Action of Vancomycin: L D D D D DDocument8 pagesMode of Action of Vancomycin: L D D D D DNolanPas encore d'évaluation
- 101jetPEI VK PDFDocument20 pages101jetPEI VK PDFsisiPas encore d'évaluation
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedD'EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedÉvaluation : 5 sur 5 étoiles5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossD'EverandThe Obesity Code: Unlocking the Secrets of Weight LossÉvaluation : 4 sur 5 étoiles4/5 (6)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDD'EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDÉvaluation : 5 sur 5 étoiles5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeD'EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeÉvaluation : 2 sur 5 étoiles2/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityD'EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityÉvaluation : 4 sur 5 étoiles4/5 (26)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionD'EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionÉvaluation : 4 sur 5 étoiles4/5 (404)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisD'EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (42)