Vous aimerez peut-être aussi
- Two Variable Data Table in ExcelDocument3 pagesTwo Variable Data Table in ExcelCandy ChungPas encore d'évaluation
- 30xl30d 26 Offset DemoDocument6 pages30xl30d 26 Offset DemoCandy ChungPas encore d'évaluation
- HandoutMsgBox PDFDocument1 pageHandoutMsgBox PDFCandy ChungPas encore d'évaluation
- Open The File in The Folder Above: Name Animal Poohsticks Score Favourite FoodDocument2 pagesOpen The File in The Folder Above: Name Animal Poohsticks Score Favourite FoodCandy ChungPas encore d'évaluation
- Name Looks Stamina Athleticism: Voting Form For JudgesDocument2 pagesName Looks Stamina Athleticism: Voting Form For JudgesCandy ChungPas encore d'évaluation
- Parts Orders Data Entry - Instructions: Input SheetDocument10 pagesParts Orders Data Entry - Instructions: Input SheetCandy ChungPas encore d'évaluation
- 30xl30d 13 Transpose DemoDocument7 pages30xl30d 13 Transpose DemoCandy ChungPas encore d'évaluation
- Milestone Chart in ExcelDocument2 pagesMilestone Chart in ExcelCandy ChungPas encore d'évaluation
- Complaint Type # of Complaints Cumulative %: Pareto Chart - Hotel ComplaintsDocument5 pagesComplaint Type # of Complaints Cumulative %: Pareto Chart - Hotel ComplaintsCandy ChungPas encore d'évaluation
- Sales Funnel ($ '000) : Download More Excel TemplatesDocument2 pagesSales Funnel ($ '000) : Download More Excel TemplatesCandy ChungPas encore d'évaluation
- Name Looks Stamina Athleticism: Voting Form For JudgesDocument2 pagesName Looks Stamina Athleticism: Voting Form For JudgesCandy ChungPas encore d'évaluation
- Poor 60% 60% Fair 15% 15% Good 15% 15% Excellent 10% 10% Value 70% 97% Target 90% 90%Document2 pagesPoor 60% 60% Fair 15% 15% Good 15% 15% Excellent 10% 10% Value 70% 97% Target 90% 90%Candy ChungPas encore d'évaluation
- Speedometer Chart in ExcelDocument2 pagesSpeedometer Chart in ExcelCandy ChungPas encore d'évaluation
- Live Time Forensics Brochuredraftv6LRDocument40 pagesLive Time Forensics Brochuredraftv6LRCandy ChungPas encore d'évaluation
- How SOIL EVIDENCE Helped Solve A Double Murder Case PDFDocument1 pageHow SOIL EVIDENCE Helped Solve A Double Murder Case PDFCandy ChungPas encore d'évaluation
- Selecting Cells in VbaDocument1 pageSelecting Cells in VbaCandy ChungPas encore d'évaluation
- Procedural Fairness, The Criminal Trial and Forensic Science and MedicineDocument36 pagesProcedural Fairness, The Criminal Trial and Forensic Science and MedicineCandy ChungPas encore d'évaluation
- How SOIL EVIDENCE Helped Solve A Double Murder CaseDocument1 pageHow SOIL EVIDENCE Helped Solve A Double Murder CaseCandy ChungPas encore d'évaluation
- Test 1: Beaumont Tower at Michigan State UniversityDocument3 pagesTest 1: Beaumont Tower at Michigan State UniversityCandy ChungPas encore d'évaluation
- Top Ten Library Excel TricksDocument3 158 pagesTop Ten Library Excel TricksCandy ChungPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
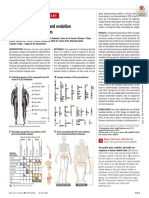
- Science - Adf8009 Genetic ArchitectureDocument13 pagesScience - Adf8009 Genetic ArchitectureHua Hidari YangPas encore d'évaluation
- Genetic TechnologyDocument36 pagesGenetic TechnologyAnggraeni Kusuma WardaniPas encore d'évaluation
- A New Tool For CRISPR-Cas13a-Based Cancer Gene TherapyDocument14 pagesA New Tool For CRISPR-Cas13a-Based Cancer Gene TherapySharon TribhuvanPas encore d'évaluation
- DNA StructureDocument4 pagesDNA StructuregabriellemichellePas encore d'évaluation
- Biological Anthropology The Natural History of Humankind 4th Edition Stanford Test BankDocument28 pagesBiological Anthropology The Natural History of Humankind 4th Edition Stanford Test Bankashleebernardgikfndpcxw100% (17)
- Solution Manual For Introduction To Biomedical Engineering 2nd Edition by DomachDocument36 pagesSolution Manual For Introduction To Biomedical Engineering 2nd Edition by Domachthionineredeyeao9j100% (44)
- Diomyka C. Damgo C31 Essay About Genetic Modification GM Technology: Is It Ethical?Document4 pagesDiomyka C. Damgo C31 Essay About Genetic Modification GM Technology: Is It Ethical?diooooomPas encore d'évaluation
- Squirrel Island ProjectDocument12 pagesSquirrel Island Projectapi-527603714Pas encore d'évaluation
- Role of Molecular Biology in Evolutionary Classification: Institute NameDocument4 pagesRole of Molecular Biology in Evolutionary Classification: Institute NameIMRAN KHANPas encore d'évaluation
- Pixl Knowledge Test Powerpoint - Aqa b2 Additional Science - Legacy 2016 and 2017Document26 pagesPixl Knowledge Test Powerpoint - Aqa b2 Additional Science - Legacy 2016 and 2017api-342297566Pas encore d'évaluation
- Polytene and Lampbrush ChromosomesDocument3 pagesPolytene and Lampbrush Chromosomesneeru.bhagatPas encore d'évaluation
- Practice Test 1Document64 pagesPractice Test 1KUSH patelPas encore d'évaluation
- Implications and Potential Role of Human Telomerase Reverse Transcriptase (Htert) in Bladder CarcinogenesisDocument7 pagesImplications and Potential Role of Human Telomerase Reverse Transcriptase (Htert) in Bladder CarcinogenesisIJAR JOURNALPas encore d'évaluation
- Miniongenetics 002Document2 pagesMiniongenetics 002api-366982317Pas encore d'évaluation
- miRDP2 Manual-V1.1.4Document13 pagesmiRDP2 Manual-V1.1.4raja sekhara reddy ravuriPas encore d'évaluation
- The CRISPR/Cas9 System and Its Applications in Crop Genome EditingDocument17 pagesThe CRISPR/Cas9 System and Its Applications in Crop Genome EditingDíaz Ramírez Brandon AlexisPas encore d'évaluation
- Human Genome ProjectDocument39 pagesHuman Genome Projectsukumar_s85% (34)
- Tumor Suppressor GenesDocument8 pagesTumor Suppressor GenesamaraadhithiyaPas encore d'évaluation
- Module 9 ACTDocument3 pagesModule 9 ACTLeighPas encore d'évaluation
- Mendelian InheritanceDocument123 pagesMendelian InheritanceAIMAN MUHAIMMIN HASNANPas encore d'évaluation
- AP Biology Lab 3 QuestionsDocument3 pagesAP Biology Lab 3 Questionsedwarddame100% (1)
- General-Biology-1 - Q1 - Week 4-1-9Document9 pagesGeneral-Biology-1 - Q1 - Week 4-1-9Paul James Dela CruzPas encore d'évaluation
- Pnas00138 0206Document5 pagesPnas00138 0206Maria VeraPas encore d'évaluation
- XyzDocument8 pagesXyzrympeikhasiPas encore d'évaluation
- Competent Cells and TransformationDocument22 pagesCompetent Cells and TransformationCarinaJongLeePas encore d'évaluation
- Meiosis Notes 2018 Fill InsDocument19 pagesMeiosis Notes 2018 Fill InsLydia LamPas encore d'évaluation
- Bioinformatics Lecture 5-9 ReviewDocument44 pagesBioinformatics Lecture 5-9 ReviewMd Saidur Rahman Kohinoor100% (3)
- Scitech 2Document2 pagesScitech 2remeceldo dagamacPas encore d'évaluation
- Fundamental Medical Science 1 Final Report (Genomic)Document14 pagesFundamental Medical Science 1 Final Report (Genomic)KesyaPas encore d'évaluation
- Hidden ConnectionsDocument15 pagesHidden ConnectionsCristina Craciun100% (2)