Vous aimerez peut-être aussi
- AACR 2022 Proceedings: Part A Online-Only and April 10D'EverandAACR 2022 Proceedings: Part A Online-Only and April 10Pas encore d'évaluation
- Indirect and Direct Methods of Gene TransferDocument4 pagesIndirect and Direct Methods of Gene TransferOrpita Das100% (2)
- 1 s2.0 S038776040900299X MainDocument6 pages1 s2.0 S038776040900299X MainLeila Schuindt MonneratPas encore d'évaluation
- Articulo 1Document11 pagesArticulo 1naPas encore d'évaluation
- Gene Expression Profiling and Pathway Analysis of Ruptured Intracranial Aneurysm (GSE36791)Document15 pagesGene Expression Profiling and Pathway Analysis of Ruptured Intracranial Aneurysm (GSE36791)Riatries SaavedraPas encore d'évaluation
- Research ArticleDocument12 pagesResearch ArticleDivya RasoiPas encore d'évaluation
- 2013 AapsjDocument8 pages2013 AapsjSCYoungPas encore d'évaluation
- Normal Hematologic and Serum Biochemical Reference Intervals For Juvenile Wild TurkeysDocument4 pagesNormal Hematologic and Serum Biochemical Reference Intervals For Juvenile Wild Turkeysweldommo2903Pas encore d'évaluation
- Digenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2Document5 pagesDigenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2SakinaPas encore d'évaluation
- Analysis of The Galactose-1-Phosphate Uridyltransferase Gene in Specimens Identified Through Newborn ScreeningDocument1 pageAnalysis of The Galactose-1-Phosphate Uridyltransferase Gene in Specimens Identified Through Newborn ScreeningboyzbanjarboyzPas encore d'évaluation
- Diabetic DisturbancesDocument18 pagesDiabetic DisturbancesshevyPas encore d'évaluation
- 1 s2.0 S0002929707624161 Main PDFDocument8 pages1 s2.0 S0002929707624161 Main PDFaris ekoPas encore d'évaluation
- 427 EndoDocument9 pages427 EndoBioq. GallegosPas encore d'évaluation
- WS 448Document3 pagesWS 448KarlaPas encore d'évaluation
- 9243 FTPDocument7 pages9243 FTPnightrider92Pas encore d'évaluation
- GBAmutations in Gaucher Type I Venezuelan Patients - Ethnic Originsand FrequenciesDocument7 pagesGBAmutations in Gaucher Type I Venezuelan Patients - Ethnic Originsand FrequenciesGilberto GomezPas encore d'évaluation
- SNP Rheumatic Arthritis Perkins2012Document4 pagesSNP Rheumatic Arthritis Perkins2012ikkePas encore d'évaluation
- Reassortment of Human Rotavirus Gene Segments Into G11 Rotavirus StrainsDocument6 pagesReassortment of Human Rotavirus Gene Segments Into G11 Rotavirus StrainsAhmed Fouad Ahmed SolimanPas encore d'évaluation
- Gene Expression Profiling in Granulosa Cells Under Levonorgestrel (2016)Document9 pagesGene Expression Profiling in Granulosa Cells Under Levonorgestrel (2016)Iris MarRoPas encore d'évaluation
- Spectrum of Mutations in Patients With Organic Acidurias From UkraineDocument10 pagesSpectrum of Mutations in Patients With Organic Acidurias From UkraineАнна ШаповаловаPas encore d'évaluation
- Choi ClinPharmacolTher 2011 PDFDocument11 pagesChoi ClinPharmacolTher 2011 PDFhuouinkyoumaPas encore d'évaluation
- Genes: The Role of Co-Deleted Genes in Neurofibromatosis Type 1 Microdeletions: An Evolutive ApproachDocument13 pagesGenes: The Role of Co-Deleted Genes in Neurofibromatosis Type 1 Microdeletions: An Evolutive ApproachMaievi FoniniPas encore d'évaluation
- Artículo 2 Ratón ObDocument5 pagesArtículo 2 Ratón ObKito TongHuiPas encore d'évaluation
- TMP 7 D6Document11 pagesTMP 7 D6FrontiersPas encore d'évaluation
- Li 2004Document8 pagesLi 2004Shukr Wesman BlbasPas encore d'évaluation
- Polymorphisms in GSTM1, GSTTI and GSTP1 and Nasopharyngeal Cancer in The East of China: A Case-Control StudyDocument4 pagesPolymorphisms in GSTM1, GSTTI and GSTP1 and Nasopharyngeal Cancer in The East of China: A Case-Control StudyontabarkerPas encore d'évaluation
- Martinez2016 PDFDocument12 pagesMartinez2016 PDFPedroFigueroaPas encore d'évaluation
- A Global Increase in 5-Hydroxymethylcytosine Levels Marks Osteoarthritic ChondrocytesDocument11 pagesA Global Increase in 5-Hydroxymethylcytosine Levels Marks Osteoarthritic Chondrocytescriters007Pas encore d'évaluation
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneDocument6 pagesDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepePas encore d'évaluation
- Variants in Nicotinic Receptors and Risk For Nicotine DependenceDocument9 pagesVariants in Nicotinic Receptors and Risk For Nicotine DependenceMuhammad AkrimPas encore d'évaluation
- 1 s2.0 S0032579119321170 MainDocument7 pages1 s2.0 S0032579119321170 MainGuilherme Costa VenturiniPas encore d'évaluation
- Blood 2008 Kroeger 1366 73Document9 pagesBlood 2008 Kroeger 1366 73Juan GomezPas encore d'évaluation
- Altered mitochondrial biology and antioxidants in neurodegenerative Fragile X disorderDocument12 pagesAltered mitochondrial biology and antioxidants in neurodegenerative Fragile X disorderPsicoterapia InfantilPas encore d'évaluation
- The Role of Germline AIP, MEN1, PRKAR1A, Cohort of Children and Adolescents With Pituitary AdenomasDocument13 pagesThe Role of Germline AIP, MEN1, PRKAR1A, Cohort of Children and Adolescents With Pituitary AdenomasBenjaMin Llacuachaqui SanchezPas encore d'évaluation
- Development of New Microsatellite DNA Markers FromDocument9 pagesDevelopment of New Microsatellite DNA Markers FromMark Louie LopezPas encore d'évaluation
- 1 s2.0 S0888754323000332 MainDocument44 pages1 s2.0 S0888754323000332 MainNour MhiriPas encore d'évaluation
- Adenosina Dra Raquel.Document13 pagesAdenosina Dra Raquel.Lalys ArualPas encore d'évaluation
- Circulating Microrna 122 in The Methionine and Choline-De Ficient Mouse Model of Non-Alcoholic SteatohepatitisDocument7 pagesCirculating Microrna 122 in The Methionine and Choline-De Ficient Mouse Model of Non-Alcoholic Steatohepatitisadamos1945Pas encore d'évaluation
- April 22021Document12 pagesApril 22021Jawaid AhmedPas encore d'évaluation
- Gen SchizopDocument11 pagesGen SchizopDewiakyuPas encore d'évaluation
- Mardinoglu 2014Document14 pagesMardinoglu 2014Julia SCPas encore d'évaluation
- Crisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Document12 pagesCrisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Saifuddin HaswarePas encore d'évaluation
- Mitochondrial Diabetes in Children Seek and You WillDocument8 pagesMitochondrial Diabetes in Children Seek and You WillsarijuicyPas encore d'évaluation
- IL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsDocument6 pagesIL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsShukr Wesman BlbasPas encore d'évaluation
- Mature and Immature Microrna Ratios in Cultured Rat Cardiomyocytes During Anoxia-ReoxygenationDocument4 pagesMature and Immature Microrna Ratios in Cultured Rat Cardiomyocytes During Anoxia-ReoxygenationTheThing81Pas encore d'évaluation
- Evidence For Diversifying Selection in Potato Virus Y and in The Coat Protein of Other PotyvirusesDocument11 pagesEvidence For Diversifying Selection in Potato Virus Y and in The Coat Protein of Other PotyvirusesTràng Hiếu NguyễnPas encore d'évaluation
- Wang 1999Document8 pagesWang 1999Sarly FebrianaPas encore d'évaluation
- Peran M Pada TikusDocument8 pagesPeran M Pada TikusShampuy ShampuyPas encore d'évaluation
- The Disruption of Profiling of Serotonergic Neurons, Results in Autism-Related BehaviorsDocument22 pagesThe Disruption of Profiling of Serotonergic Neurons, Results in Autism-Related BehaviorsAngelinni Taglioni StangePas encore d'évaluation
- Treg 5Document4 pagesTreg 5AlisPas encore d'évaluation
- Allele Frequencies of Six MiniSTR Loci in The Population of Northern PortugalDocument3 pagesAllele Frequencies of Six MiniSTR Loci in The Population of Northern PortugalspanishvcuPas encore d'évaluation
- BypassDocument5 pagesBypassGeorgina HernandezPas encore d'évaluation
- Malay DnaDocument9 pagesMalay DnaEffandi Abu BakarPas encore d'évaluation
- 10 - Chapter 4Document10 pages10 - Chapter 4The FrequencyPas encore d'évaluation
- Pyruvate Kinase Deficiency and Malaria: Brief ReportDocument6 pagesPyruvate Kinase Deficiency and Malaria: Brief ReportErlangga Perwira NegaraPas encore d'évaluation
- A New LC MS MS Method For Quantification of Gangliosides in Human Plasma PDFDocument32 pagesA New LC MS MS Method For Quantification of Gangliosides in Human Plasma PDFRamona Neka TamoPas encore d'évaluation
- tmp1329 TMPDocument11 pagestmp1329 TMPFrontiersPas encore d'évaluation
- Human population genetics and phenotype dataDocument6 pagesHuman population genetics and phenotype dataquique ddmPas encore d'évaluation
- Assays: Protein: Bicinchoninic Acid (BCA) MethodDocument2 pagesAssays: Protein: Bicinchoninic Acid (BCA) MethodNataraj NagarajanPas encore d'évaluation
- Hallmark of CancerDocument14 pagesHallmark of CanceriinsabatiniPas encore d'évaluation
- Cellular and Molecular Basis of InheritanceDocument71 pagesCellular and Molecular Basis of InheritancevalikoPas encore d'évaluation
- 1.4. Cell StructureDocument4 pages1.4. Cell StructuresamPas encore d'évaluation
- Molecular Basis of InheritanceDocument52 pagesMolecular Basis of InheritancezoologyPas encore d'évaluation
- Cell Revision WorksheetDocument3 pagesCell Revision WorksheetCally Chew100% (3)
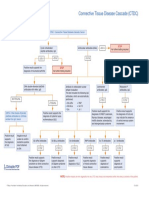
- Connective Tissue Disease CascadeDocument1 pageConnective Tissue Disease CascadeYustina BubnovaPas encore d'évaluation
- 1.metab Nukleotida 1Document66 pages1.metab Nukleotida 1DifaniaPas encore d'évaluation
- Lehninger Ch7 CHOSDocument31 pagesLehninger Ch7 CHOSIsmael ChablePas encore d'évaluation
- People To Observe and Describe Microorganisms Generation: LivingDocument7 pagesPeople To Observe and Describe Microorganisms Generation: LivingSabnin SoroniPas encore d'évaluation
- Activity For Grade 9 RubbyDocument5 pagesActivity For Grade 9 RubbyJocelyn Petallar BalasuelaPas encore d'évaluation
- As Biology Assignment 2 Food and HealthDocument11 pagesAs Biology Assignment 2 Food and HealthmuhajireenPas encore d'évaluation
- AS LEVEL BIOLOGY Paper 1 EnzymesDocument54 pagesAS LEVEL BIOLOGY Paper 1 EnzymesADEEL AHMADPas encore d'évaluation
- Adapterama Iii: Quadruple-Indexed, Triple-Enzyme Radseq Libraries For About $1usd Per Sample (3rad)Document35 pagesAdapterama Iii: Quadruple-Indexed, Triple-Enzyme Radseq Libraries For About $1usd Per Sample (3rad)OanaPas encore d'évaluation
- Big Picture: Genes, Genomes and HealthDocument16 pagesBig Picture: Genes, Genomes and HealthWellcome Trust100% (2)
- 1612091342Document290 pages1612091342mgerke67576Pas encore d'évaluation
- Carbohydrates: Disaccharides and PolysaccharidesDocument3 pagesCarbohydrates: Disaccharides and PolysaccharidesTherese Vince J. LazosPas encore d'évaluation
- Inheritance & Mutation PatternsDocument25 pagesInheritance & Mutation PatternsJennifer Dixon100% (1)
- Blood Bank. Harmening Chapter 2 Review QuestionsDocument2 pagesBlood Bank. Harmening Chapter 2 Review QuestionsMerry GracePas encore d'évaluation
- Lahat NG Sagot Sa GBIO Andito NaDocument36 pagesLahat NG Sagot Sa GBIO Andito NaNix Prieto100% (1)
- 35 1419 1 PBDocument7 pages35 1419 1 PBKhaerul FadlyPas encore d'évaluation
- Grade 9 DNADocument19 pagesGrade 9 DNARitchwel FAJARDO100% (1)
- Gen & Genom TumbuhanDocument33 pagesGen & Genom Tumbuhanmonocotil9879Pas encore d'évaluation
- Human Genetics: From DNA to ProteinsDocument28 pagesHuman Genetics: From DNA to ProteinsjasPas encore d'évaluation
- Yeast Autolysis in Sparkling Wine - A Review, SUBDocument9 pagesYeast Autolysis in Sparkling Wine - A Review, SUBoscardannstrom5431Pas encore d'évaluation
- Pathophysiology of Type 1 Diabetes MellitusDocument13 pagesPathophysiology of Type 1 Diabetes MellitusPelle MozzaPas encore d'évaluation
- CellsDocument31 pagesCellsPranav ShindePas encore d'évaluation
- Genetic TechnologyDocument4 pagesGenetic TechnologyJelin VergaraPas encore d'évaluation
- Immunoglobulins: Prof - Dr.Gülden Burçak 2020-2021Document25 pagesImmunoglobulins: Prof - Dr.Gülden Burçak 2020-2021Marwa AliPas encore d'évaluation