Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- 4 - Odd Hours - Dean KoontzDocument261 pages4 - Odd Hours - Dean KoontzJustinSnow80% (5)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Dr. Berg's Favorite HEALTHY JUNK FOODS & Other AlternativesDocument23 pagesDr. Berg's Favorite HEALTHY JUNK FOODS & Other Alternativesprashant_padte100% (8)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Project Report On Air FilterDocument5 pagesProject Report On Air FilterEIRI Board of Consultants and PublishersPas encore d'évaluation
- SCAT Chart - Systematic Cause Analysis Technique - SCAT ChartDocument6 pagesSCAT Chart - Systematic Cause Analysis Technique - SCAT ChartSalman Alfarisi100% (1)
- HBSE-Mock ExamDocument3 pagesHBSE-Mock ExamAnnePas encore d'évaluation
- Foundations in Microbiology: Elements of Microbial Nutrition, Ecology, and Growth TalaroDocument47 pagesFoundations in Microbiology: Elements of Microbial Nutrition, Ecology, and Growth Talaroanil_hodPas encore d'évaluation
- Movie Ethics ReviewDocument4 pagesMovie Ethics ReviewpearlydawnPas encore d'évaluation
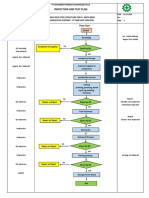
- Inspection and Test Plan: Flow Chart Start IncomingDocument1 pageInspection and Test Plan: Flow Chart Start IncomingSinden AyuPas encore d'évaluation
- Catalyst Loading ProcedureDocument3 pagesCatalyst Loading ProcedureNaresh SamalaPas encore d'évaluation
- Ogino 2008 New Middle Pleistocene Galictini (Mustelidae, Carnivora)Document9 pagesOgino 2008 New Middle Pleistocene Galictini (Mustelidae, Carnivora)Ghaier KazmiPas encore d'évaluation
- Dong Et Al. 2010 Age and Paleoenvironment of Xiaogushan Fauna at Haicheng, Liaoning ProvinceDocument6 pagesDong Et Al. 2010 Age and Paleoenvironment of Xiaogushan Fauna at Haicheng, Liaoning ProvinceGhaier KazmiPas encore d'évaluation
- Large Mustelid Species from Netherlands SiteDocument6 pagesLarge Mustelid Species from Netherlands SiteGhaier KazmiPas encore d'évaluation
- Hall 1930 A Bassarisk and A New Mustelid From The Later Tertiary of CaliforniaDocument5 pagesHall 1930 A Bassarisk and A New Mustelid From The Later Tertiary of CaliforniaGhaier KazmiPas encore d'évaluation
- Hall 1935 A New Mustelid Genus From The Pliocene of CaliforniaDocument3 pagesHall 1935 A New Mustelid Genus From The Pliocene of CaliforniaGhaier KazmiPas encore d'évaluation
- Body mass estimation of extinct "beardogDocument22 pagesBody mass estimation of extinct "beardogGhaier KazmiPas encore d'évaluation
- Geraads 2016 Pleistocene Carnivora (Mammalia) From Tighennif (Ternifine), AlgeriaDocument45 pagesGeraads 2016 Pleistocene Carnivora (Mammalia) From Tighennif (Ternifine), AlgeriaGhaier KazmiPas encore d'évaluation
- Tehreef From Faizan Ul Rehman Vol 1-2Document27 pagesTehreef From Faizan Ul Rehman Vol 1-2Ghaier KazmiPas encore d'évaluation
- Wish To Join BiotechDocument1 pageWish To Join BiotechGhaier KazmiPas encore d'évaluation
- Study of Karyotypes (Giant Polytene Chromosomes) in Drosophila MelanogasterDocument3 pagesStudy of Karyotypes (Giant Polytene Chromosomes) in Drosophila MelanogasterGhaier KazmiPas encore d'évaluation
- Some New Fossils of Protanancus Chinjiensis (Proboscidea, Mammalia) From Middle Miocene of PakistanDocument5 pagesSome New Fossils of Protanancus Chinjiensis (Proboscidea, Mammalia) From Middle Miocene of PakistanGhaier KazmiPas encore d'évaluation
- Khan & Sarwar 1993 A New Fossil Giraffid Species Bramatherium Progressus N. Sp. From Siwalik Formations of PakistanDocument9 pagesKhan & Sarwar 1993 A New Fossil Giraffid Species Bramatherium Progressus N. Sp. From Siwalik Formations of PakistanGhaier KazmiPas encore d'évaluation
- Study of Cytochemical Detection of DNA in Protozoa or Avian Blood CelDocument3 pagesStudy of Cytochemical Detection of DNA in Protozoa or Avian Blood CelGhaier Kazmi0% (1)
- Khan Et Al. 2002 A New Large Bramatherium Giraffe Bramatherium Giganteus Sp. NovDocument5 pagesKhan Et Al. 2002 A New Large Bramatherium Giraffe Bramatherium Giganteus Sp. NovGhaier KazmiPas encore d'évaluation
- Cambridge Books OnlineDocument5 pagesCambridge Books OnlineGhaier KazmiPas encore d'évaluation
- McRae 1990 Paleomagnetic Isochrons, Unsteadiness, and Non-Uniformity of Sedimentation in Miocene FluvialDocument24 pagesMcRae 1990 Paleomagnetic Isochrons, Unsteadiness, and Non-Uniformity of Sedimentation in Miocene FluvialGhaier KazmiPas encore d'évaluation
- Abstract PublishedDocument2 pagesAbstract PublishedGhaier KazmiPas encore d'évaluation
- CBO9780511793844.002 Fossil Primates - Primate TaxonomyDocument23 pagesCBO9780511793844.002 Fossil Primates - Primate TaxonomyGhaier KazmiPas encore d'évaluation
- Geological Time ScaleDocument2 pagesGeological Time ScaleGhaier KazmiPas encore d'évaluation
- Difference Between Native, Endemic, Introduced and Invasive SpeciesDocument3 pagesDifference Between Native, Endemic, Introduced and Invasive SpeciesGhaier KazmiPas encore d'évaluation
- CBO9780511793844.005 Fossil Primates - The World of The PastDocument8 pagesCBO9780511793844.005 Fossil Primates - The World of The PastGhaier KazmiPas encore d'évaluation
- CBO9780511793844.006 Fossil Primates - The Lifeways of Extinct AnimalsDocument25 pagesCBO9780511793844.006 Fossil Primates - The Lifeways of Extinct AnimalsGhaier KazmiPas encore d'évaluation
- Palynomorph Extraction From Peat, Lignite and CoalDocument5 pagesPalynomorph Extraction From Peat, Lignite and CoalGhaier KazmiPas encore d'évaluation
- CBO9780511793844.008 Fossil Primates - Primate OriginsDocument18 pagesCBO9780511793844.008 Fossil Primates - Primate OriginsGhaier KazmiPas encore d'évaluation
- CBO9780511793844.007 Fossil Primates - Evolutionary Processes and The Pattern of Primate EvolutionDocument14 pagesCBO9780511793844.007 Fossil Primates - Evolutionary Processes and The Pattern of Primate EvolutionGhaier KazmiPas encore d'évaluation
- CBO9780511793844.005 Fossil Primates - The World of The PastDocument8 pagesCBO9780511793844.005 Fossil Primates - The World of The PastGhaier KazmiPas encore d'évaluation
- Schneider - Cptg010 en (Print)Document16 pagesSchneider - Cptg010 en (Print)el_koptan00857693Pas encore d'évaluation
- Sunflower Herbicide ChartDocument2 pagesSunflower Herbicide ChartpapucicaPas encore d'évaluation
- Rajasthan PET Chemistry 1997 Exam with Detailed SolutionsDocument34 pagesRajasthan PET Chemistry 1997 Exam with Detailed SolutionsYumit MorwalPas encore d'évaluation
- CASR Part 830 Amdt. 2 - Notification & Reporting of Aircraft Accidents, Incidents, or Overdue Acft & Investigation OCRDocument17 pagesCASR Part 830 Amdt. 2 - Notification & Reporting of Aircraft Accidents, Incidents, or Overdue Acft & Investigation OCRHarry NuryantoPas encore d'évaluation
- Corn Pulao Recipe With Sweet CornDocument2 pagesCorn Pulao Recipe With Sweet CornSudharshanPas encore d'évaluation
- Report Experiment 5 STK1211Document9 pagesReport Experiment 5 STK1211NurAkila Mohd YasirPas encore d'évaluation
- Rice Research: Open Access: Black Rice Cultivation and Forming Practices: Success Story of Indian FarmersDocument2 pagesRice Research: Open Access: Black Rice Cultivation and Forming Practices: Success Story of Indian Farmersapi-420356823Pas encore d'évaluation
- Temporomandibular Joint SyndromeDocument11 pagesTemporomandibular Joint SyndromeRahma RahmaPas encore d'évaluation
- Naskah PublikasiDocument14 pagesNaskah PublikasiNirmala malaPas encore d'évaluation
- Radiol 2020201473Document37 pagesRadiol 2020201473M Victoria SalazarPas encore d'évaluation
- Rundingan Perdagangan Antara Malaysia Dan Indonesia Di Wisma Putra, Kuala Lumpur 1967Document15 pagesRundingan Perdagangan Antara Malaysia Dan Indonesia Di Wisma Putra, Kuala Lumpur 1967nixyingboPas encore d'évaluation
- Somali Guideline of InvestorsDocument9 pagesSomali Guideline of InvestorsABDULLAHI HAGAR FARAH HERSI STUDENTPas encore d'évaluation
- Refresher 2 (Pipe)Document12 pagesRefresher 2 (Pipe)Kira YagamiPas encore d'évaluation
- Byk-A 525 enDocument2 pagesByk-A 525 enさいとはちこPas encore d'évaluation
- 1324 1624 1824 Owners Manual 6 4 08Document64 pages1324 1624 1824 Owners Manual 6 4 08Miguel LopPas encore d'évaluation
- EFPSDocument8 pagesEFPSBryan Joshua VillarPas encore d'évaluation
- Volume 1Document168 pagesVolume 1lalitendu jenaPas encore d'évaluation
- Baileys in 2009: Case Study Reference No 509-050-1Document17 pagesBaileys in 2009: Case Study Reference No 509-050-1Ernesto KulasinPas encore d'évaluation
- Chemistry CHM 1311C 2012 Test 1 BlankDocument6 pagesChemistry CHM 1311C 2012 Test 1 BlankSimon HagosPas encore d'évaluation
- INFORSHT Produktkatalog en Web 03.22Document13 pagesINFORSHT Produktkatalog en Web 03.22lolPas encore d'évaluation
- Calculation of Lightning and Switching Overvoltages Transferred Through Power TransformerDocument9 pagesCalculation of Lightning and Switching Overvoltages Transferred Through Power TransformerBožidar Filipović-GrčićPas encore d'évaluation
- Chin Cup Therapy An Effective Tool For The Correction of Class III Malocclusion in Mixed and Late Deciduous DentitionsDocument6 pagesChin Cup Therapy An Effective Tool For The Correction of Class III Malocclusion in Mixed and Late Deciduous Dentitionschic organizerPas encore d'évaluation