Vous aimerez peut-être aussi
- New Biological Anthropology PDFDocument11 pagesNew Biological Anthropology PDFRubén García DíazPas encore d'évaluation
- Christianity As Mystical FactDocument272 pagesChristianity As Mystical Factqwertyp100% (2)
- Soft Tissue Mobilization: SP Ecial Con Si Der Ation SDocument42 pagesSoft Tissue Mobilization: SP Ecial Con Si Der Ation SVizaPas encore d'évaluation
- New Senior Secondary MASTERING BIOLOGY OXFORD Mock - Bio - Set6 - eDocument45 pagesNew Senior Secondary MASTERING BIOLOGY OXFORD Mock - Bio - Set6 - eChun Kit LauPas encore d'évaluation
- DRILLSDocument3 pagesDRILLSDeepak Prakash Jaya100% (4)
- DansdynamicwarmupspeeddayDocument2 pagesDansdynamicwarmupspeedday44dwymanPas encore d'évaluation
- Alexandra David-Neel - Magic and Mystery in Tibet (105p) (Inua) PDFDocument104 pagesAlexandra David-Neel - Magic and Mystery in Tibet (105p) (Inua) PDFAngelo Karageorgos100% (1)
- Stretching: Muscle Strength For SpeedDocument4 pagesStretching: Muscle Strength For SpeedGerardClarkePas encore d'évaluation
- Weight Training Medium Interval Training Medium Interval Training Weight Training Rugby Specific Skill CircuitDocument1 pageWeight Training Medium Interval Training Medium Interval Training Weight Training Rugby Specific Skill CircuitJess FarrellPas encore d'évaluation
- Forever Fat Loss - Cs PDFDocument263 pagesForever Fat Loss - Cs PDFDavid CieloPas encore d'évaluation
- Nutrition For The AthleteDocument11 pagesNutrition For The AthletewenghuatPas encore d'évaluation
- Lower Extremity Stretching ProtocolDocument3 pagesLower Extremity Stretching ProtocolHello RainbowPas encore d'évaluation
- NLP - The Magic You Can Perform With Reframing (By Michael Hall PH.D)Document4 pagesNLP - The Magic You Can Perform With Reframing (By Michael Hall PH.D)pinkyoyo100% (1)
- The Complete Jason Blaha Ice Cream Fitness 5x5 NoviceProgramDocument11 pagesThe Complete Jason Blaha Ice Cream Fitness 5x5 NoviceProgramgastaoPas encore d'évaluation
- Curriculum GuidebookDocument25 pagesCurriculum GuidebookArtemisica GPas encore d'évaluation
- Get Max Vertical Hot HandlesDocument59 pagesGet Max Vertical Hot HandlesWilliam MosqueraPas encore d'évaluation
- Rehab U ShoulderfixDocument11 pagesRehab U ShoulderfixCharles MitchellPas encore d'évaluation
- Gnostic Study of The Soul PDFDocument5 pagesGnostic Study of The Soul PDFAngelo KarageorgosPas encore d'évaluation
- The Nourished Kitchen Guide To Grain-Free Baking, Treats and SweetsDocument76 pagesThe Nourished Kitchen Guide To Grain-Free Baking, Treats and SweetsAngelo Karageorgos100% (1)
- Nutritional PlanDocument20 pagesNutritional Planapi-259457599Pas encore d'évaluation
- Lumbar Core StrenghtDocument8 pagesLumbar Core StrenghtMattia BressanPas encore d'évaluation
- Cape CrossFit Athletic Skill LevelsDocument1 pageCape CrossFit Athletic Skill LevelsIsaac Gordo FloresPas encore d'évaluation
- Joey Bergles - Lifting Wks 5-8 (Minimal Equipment)Document4 pagesJoey Bergles - Lifting Wks 5-8 (Minimal Equipment)George CarrascoPas encore d'évaluation
- CBB - Program Phase 1Document4 pagesCBB - Program Phase 1Titus BaldwinPas encore d'évaluation
- Rings 1 Getting Started GuideDocument13 pagesRings 1 Getting Started GuidedraguPas encore d'évaluation
- Eat Up Raise Your Game: 100 easy, nutritious recipes to help you perform better on exercise days and rest daysD'EverandEat Up Raise Your Game: 100 easy, nutritious recipes to help you perform better on exercise days and rest daysPas encore d'évaluation
- Form 2 Chapter 2Document7 pagesForm 2 Chapter 2naza977579% (19)
- Hip Mobility Sequence PDFDocument1 pageHip Mobility Sequence PDFgaurnityanandaPas encore d'évaluation
- Run & Hurdle Technique Technique & Drills SpottingDocument3 pagesRun & Hurdle Technique Technique & Drills Spottingcacamerba90Pas encore d'évaluation
- Pullups Workout ProgramDocument6 pagesPullups Workout ProgramZe Luis TerrassoPas encore d'évaluation
- ACL Reconstruction Rehabilitation GuidelinesDocument4 pagesACL Reconstruction Rehabilitation Guidelinesfia0711Pas encore d'évaluation
- Bill Starr - A Strong StartDocument6 pagesBill Starr - A Strong StartCoco BungbingPas encore d'évaluation
- 10 Secrets in 10 YearsDocument15 pages10 Secrets in 10 YearsasicsPas encore d'évaluation
- The Spine - A StructuralDocument2 pagesThe Spine - A Structuralcraigvh89Pas encore d'évaluation
- Baremetrics - Growing A Million Dollar Company by Josh PigfordDocument216 pagesBaremetrics - Growing A Million Dollar Company by Josh PigfordGonzalo ZariagaPas encore d'évaluation
- How To Improve Your Vertical LeapDocument12 pagesHow To Improve Your Vertical Leapskilzace100% (1)
- BellyOff-Jumpstart Warm UpDocument8 pagesBellyOff-Jumpstart Warm UpocforuPas encore d'évaluation
- Crandall, B.C. (Ed.) - NanotechnologyDocument216 pagesCrandall, B.C. (Ed.) - Nanotechnologyjacobi84Pas encore d'évaluation
- Build More On The FloorDocument1 pageBuild More On The FloorČedomir Žarković100% (1)
- Metabolic Conditioning Methods ChartDocument1 pageMetabolic Conditioning Methods ChartKarlosPas encore d'évaluation
- Interview With Master Samael Aun Weor 1977Document19 pagesInterview With Master Samael Aun Weor 1977Donald William100% (1)
- Why Humans Have SexDocument31 pagesWhy Humans Have Sexpeter_martin93350% (1)
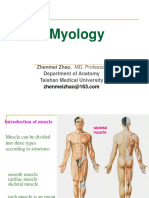
- Myology: Zhenmei Zhao, MD, Professor Department of Anatomy Taishan Medical UniversityDocument123 pagesMyology: Zhenmei Zhao, MD, Professor Department of Anatomy Taishan Medical UniversityAkshit SetiaPas encore d'évaluation
- MC 20Document9 pagesMC 20Meu BroPas encore d'évaluation
- Oxygen Cost of Kettlebell SwingDocument3 pagesOxygen Cost of Kettlebell SwingDamiano MolinaroPas encore d'évaluation
- Ancestral Health Symposium 2011 ProgramDocument3 pagesAncestral Health Symposium 2011 ProgramBryan CastañedaPas encore d'évaluation
- CFES PTRM SamplePgsDocument70 pagesCFES PTRM SamplePgsIvo LeitePas encore d'évaluation
- Core Stability Rippetoe PDFDocument4 pagesCore Stability Rippetoe PDFSylvia GracePas encore d'évaluation
- Chap 8 PDFDocument5 pagesChap 8 PDFQuốc HuyPas encore d'évaluation
- Physical Agility Test Preparation and Safety: How To Prepare For The PATDocument9 pagesPhysical Agility Test Preparation and Safety: How To Prepare For The PATJmFormalesPas encore d'évaluation
- W336 LEARNING TASK No. 1.2 PARAGESDocument2 pagesW336 LEARNING TASK No. 1.2 PARAGESJeric ParagesPas encore d'évaluation
- Liebenson - Functional Training Handbook ReviewDocument3 pagesLiebenson - Functional Training Handbook ReviewJoaoPncs SilvaPas encore d'évaluation
- Trunk Stability by Calvin Morriss, PHD.: What Exactly Is Trunk Stability For A Rugby Player?Document9 pagesTrunk Stability by Calvin Morriss, PHD.: What Exactly Is Trunk Stability For A Rugby Player?Justice Man100% (1)
- Bench Press SyndromeDocument4 pagesBench Press SyndromeAbdallah Maroun NajemPas encore d'évaluation
- Jump Rope TrainingDocument4 pagesJump Rope TrainingpapircsillagPas encore d'évaluation
- Chapter 11 - RecoveryDocument20 pagesChapter 11 - RecoveryDDVPas encore d'évaluation
- Tri-Phasic Epigenetic Model of Self-AcceptanceDocument1 pageTri-Phasic Epigenetic Model of Self-AcceptanceLucPas encore d'évaluation
- Lean Muscle ProgramDocument2 pagesLean Muscle ProgramtonypodmorePas encore d'évaluation
- Batman Workout! PDFDocument3 pagesBatman Workout! PDFGiamma GiammaPas encore d'évaluation
- Velocity TrainingDocument8 pagesVelocity Trainingjkent0150Pas encore d'évaluation
- The Vert Specific Conditioning Guide : How Hard Can You Go and For How Long?.Document20 pagesThe Vert Specific Conditioning Guide : How Hard Can You Go and For How Long?.Luis Ferdinand Dacera-Gabronino Gamponia-NonanPas encore d'évaluation
- Squat Glute Challenge - en PDFDocument1 pageSquat Glute Challenge - en PDFArpit LadolePas encore d'évaluation
- Mechanical Demands of Kettlebell Swing ExerciseDocument25 pagesMechanical Demands of Kettlebell Swing ExerciseDamiano MolinaroPas encore d'évaluation
- Running Head: An Analysis of The Squat 1Document7 pagesRunning Head: An Analysis of The Squat 1Karanja KinyanjuiPas encore d'évaluation
- 2.2 - All Worksheets - Homework Sheets - Extension SheetsDocument20 pages2.2 - All Worksheets - Homework Sheets - Extension SheetsloginePas encore d'évaluation
- Peak Fitness WorkoutDocument6 pagesPeak Fitness WorkoutIsael AlemanPas encore d'évaluation
- 5 Nutrition Mistakes Athletes MakeDocument3 pages5 Nutrition Mistakes Athletes MakeScott McLanePas encore d'évaluation
- How Much Can You LiftDocument6 pagesHow Much Can You LiftaboubakereldjazairiPas encore d'évaluation
- The DeadliftDocument8 pagesThe DeadliftNilgunPas encore d'évaluation
- Molecular and Cellular Aspects of Muscle Function: Proceedings of the 28th International Congress of Physiological Sciences Budapest 1980, (including the proceedings of the satellite symposium on Membrane Control of Skeletal Muscle Function)D'EverandMolecular and Cellular Aspects of Muscle Function: Proceedings of the 28th International Congress of Physiological Sciences Budapest 1980, (including the proceedings of the satellite symposium on Membrane Control of Skeletal Muscle Function)E. VargaPas encore d'évaluation
- Aphorisms Nassim TalebDocument5 pagesAphorisms Nassim TalebAngelo KarageorgosPas encore d'évaluation
- Coin Collecting Basics: Your Own Coin Collecting KitDocument2 pagesCoin Collecting Basics: Your Own Coin Collecting KitAngelo KarageorgosPas encore d'évaluation
- Serial Monogamy As Polygyny or Polyandry?Document22 pagesSerial Monogamy As Polygyny or Polyandry?Angelo KarageorgosPas encore d'évaluation
- International Society of Sports Nutrition Position Stand Diets and Body CompositionDocument19 pagesInternational Society of Sports Nutrition Position Stand Diets and Body CompositionLuis Ricardo Hernandez GarciaPas encore d'évaluation
- Attractive Women Want It All. Good Genes, Economic Investment, Parenting Proclivities, and Emotional Commitment PDFDocument13 pagesAttractive Women Want It All. Good Genes, Economic Investment, Parenting Proclivities, and Emotional Commitment PDFAngelo KarageorgosPas encore d'évaluation
- International Society of Sports Nutrition Position Stand Diets and Body CompositionDocument19 pagesInternational Society of Sports Nutrition Position Stand Diets and Body CompositionLuis Ricardo Hernandez GarciaPas encore d'évaluation
- 7 Steps To Better Grain-Free BakingDocument29 pages7 Steps To Better Grain-Free BakingMarmotiniPas encore d'évaluation
- BIOSC011 38268 HumanBio Neary S15Document10 pagesBIOSC011 38268 HumanBio Neary S15Emily PhanPas encore d'évaluation
- Jurnal Lime JuiceDocument9 pagesJurnal Lime JuicenanaazzahraPas encore d'évaluation
- Biology IGCSE Paper 4Document20 pagesBiology IGCSE Paper 4veronicaPas encore d'évaluation
- Nitrate Reduction in Sulfate Reducing BacteriaDocument10 pagesNitrate Reduction in Sulfate Reducing BacteriaCatalinaManjarresPas encore d'évaluation
- EndocnidozoaDocument6 pagesEndocnidozoaKevin Rucci100% (1)
- ChymosinDocument242 pagesChymosinvbortizlPas encore d'évaluation
- Student Exploration: Digestive SystemDocument4 pagesStudent Exploration: Digestive SystemNO POGGEGRPas encore d'évaluation
- Marocmolecule Review Worksheet - Answer KeyDocument3 pagesMarocmolecule Review Worksheet - Answer KeyBecka RodriguezPas encore d'évaluation
- Worksheet FermentationDocument5 pagesWorksheet FermentationRyan De AlloPas encore d'évaluation
- 108 PDFDocument7 pages108 PDFpuspa sariPas encore d'évaluation
- Chapter 23 RQDocument4 pagesChapter 23 RQMia MistypuffPas encore d'évaluation
- Classification Notes PDFDocument20 pagesClassification Notes PDFBobPas encore d'évaluation
- Scooby's Suggested Solutions For The 2014 VCE Biology Exam - Short AnswerDocument6 pagesScooby's Suggested Solutions For The 2014 VCE Biology Exam - Short AnswerRite10210Pas encore d'évaluation
- Nutritional Aspects of Exercise and Sport Performance PlannerDocument2 pagesNutritional Aspects of Exercise and Sport Performance PlannerUSman TArarPas encore d'évaluation
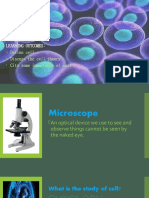
- Unit 1:: Learning OutcomesDocument15 pagesUnit 1:: Learning OutcomesJhenny Lyn CamanoPas encore d'évaluation
- Introduction To Medical Microbiology 2023Document93 pagesIntroduction To Medical Microbiology 2023Samuel fikaduPas encore d'évaluation
- Plant Kingdom PowernotesDocument3 pagesPlant Kingdom Powernotessheikhanzar643Pas encore d'évaluation
- Lesson 4Document7 pagesLesson 4Jay Lester UrsolinoPas encore d'évaluation
- Devoir de Synthèse N°2 2012 2013 (Hajji Abderrazek)Document4 pagesDevoir de Synthèse N°2 2012 2013 (Hajji Abderrazek)NihedPas encore d'évaluation
- 3 Agarose Gel Electrophoresis & Ames TestDocument4 pages3 Agarose Gel Electrophoresis & Ames TestNurulPas encore d'évaluation
- NCM 112 MW NotesDocument6 pagesNCM 112 MW NotesJames Kristopher RebayaPas encore d'évaluation
- 1 TRGSCHD NIPHM 2012 13Document12 pages1 TRGSCHD NIPHM 2012 13Vikas NikharangePas encore d'évaluation
- The Importance of A Balanced DietDocument4 pagesThe Importance of A Balanced DietdimdamflyPas encore d'évaluation
- Hope - 1 Grade 11: Exercise For FitnessDocument11 pagesHope - 1 Grade 11: Exercise For FitnessMark Johnson San JuanPas encore d'évaluation
- BCC - DNA Replication in Prokaryotes Lecture 20Document31 pagesBCC - DNA Replication in Prokaryotes Lecture 20AmyPas encore d'évaluation
- Grupo H. Aspergillus Fumigatus and Aspergillosis From Basics To ClinicsDocument51 pagesGrupo H. Aspergillus Fumigatus and Aspergillosis From Basics To Clinicsjaver andres hernandezveraPas encore d'évaluation