Vous aimerez peut-être aussi
- Chicago GridDocument1 pageChicago GridMichael PearsonPas encore d'évaluation
- Dynamics and Structure in Metallic Supercooled LiDocument119 pagesDynamics and Structure in Metallic Supercooled LiMichael PearsonPas encore d'évaluation
- Crystallization of Fat in and Outside Milk Fat Globules Uden Artikler PABUDocument90 pagesCrystallization of Fat in and Outside Milk Fat Globules Uden Artikler PABUMichael PearsonPas encore d'évaluation
- Brakke - Surface EvolverDocument25 pagesBrakke - Surface EvolverMichael PearsonPas encore d'évaluation
- What Is Quantum Cognition, and How Is It Applied To PsychologyDocument7 pagesWhat Is Quantum Cognition, and How Is It Applied To PsychologyMichael PearsonPas encore d'évaluation
- Molecular Computing Does DNA ComputeDocument4 pagesMolecular Computing Does DNA ComputeMichael PearsonPas encore d'évaluation
- Chronicle of Higher Ed Ground Zero For Unionization Nov 23 2015Document7 pagesChronicle of Higher Ed Ground Zero For Unionization Nov 23 2015Michael PearsonPas encore d'évaluation
- Advanced Operating Systems Denning Brown1984Document18 pagesAdvanced Operating Systems Denning Brown1984Michael PearsonPas encore d'évaluation
- LyapunovDocument32 pagesLyapunovMichael PearsonPas encore d'évaluation
- Ramparts CIA FrontDocument10 pagesRamparts CIA FrontMichael PearsonPas encore d'évaluation
- Refining and Processing SugarDocument4 pagesRefining and Processing SugarRocío CapuñayPas encore d'évaluation
- Steiner Trees and Spanning Trees in Six-Pin Soap FilmsDocument17 pagesSteiner Trees and Spanning Trees in Six-Pin Soap FilmsMichael PearsonPas encore d'évaluation
- Entangled States of Trapped Atomic Blatt 2008Document8 pagesEntangled States of Trapped Atomic Blatt 2008Oliver BarterPas encore d'évaluation
- Relative Entropy in Quantum Information TheoryDocument36 pagesRelative Entropy in Quantum Information TheoryMichael Pearson100% (1)
- Worlds Muslims Religion Politics Society Full ReportDocument226 pagesWorlds Muslims Religion Politics Society Full ReportEduardo RossiPas encore d'évaluation
- Elisabeth Verhoeven Experiential Constructions in Yucatec Maya A Typologically Based Analysis of A Functional Domain in A Mayan Language Studies in Language Companion SDocument405 pagesElisabeth Verhoeven Experiential Constructions in Yucatec Maya A Typologically Based Analysis of A Functional Domain in A Mayan Language Studies in Language Companion SMichael Pearson100% (1)
- Tracking The Origins of A State Terror NetworkDocument23 pagesTracking The Origins of A State Terror NetworkMichael PearsonPas encore d'évaluation
- Quantum Computation and Shor's Factoring AlgorithmDocument21 pagesQuantum Computation and Shor's Factoring AlgorithmMichael PearsonPas encore d'évaluation
- Electrochemical Corrosion of Unalloyed Copper in Chloride Media - A Critical ReviewDocument27 pagesElectrochemical Corrosion of Unalloyed Copper in Chloride Media - A Critical ReviewMichael PearsonPas encore d'évaluation
- Electrically Induced Morphological Instabilities in Free Dendrite GrowthDocument4 pagesElectrically Induced Morphological Instabilities in Free Dendrite GrowthMichael PearsonPas encore d'évaluation
- Scale Relativity and Schrödinger's EquationDocument17 pagesScale Relativity and Schrödinger's EquationMichael PearsonPas encore d'évaluation
- 5.time Symmetry Breaking, DualityDocument19 pages5.time Symmetry Breaking, Dualitypipul36Pas encore d'évaluation
- 10 Big Problems with Mandatory E-Verify SystemDocument13 pages10 Big Problems with Mandatory E-Verify SystemMichael PearsonPas encore d'évaluation
- Foundations of Algebraic GeometryDocument764 pagesFoundations of Algebraic GeometryMichael Pearson100% (2)
- Microfluidics For DNA AnalysisDocument16 pagesMicrofluidics For DNA AnalysisMichael PearsonPas encore d'évaluation
- Valuation of Ruil Forests Ecosystem Services in Central Chile With The Choice Experiment MethodDocument30 pagesValuation of Ruil Forests Ecosystem Services in Central Chile With The Choice Experiment MethodMichael PearsonPas encore d'évaluation
- United States: (12) Patent Application Publication (10) Pub. No.: US 2001/0044775 A1Document59 pagesUnited States: (12) Patent Application Publication (10) Pub. No.: US 2001/0044775 A1Michael PearsonPas encore d'évaluation
- United States: (12) Patent Application Publication (10) Pub. No.: US 2014/0218233 A1Document18 pagesUnited States: (12) Patent Application Publication (10) Pub. No.: US 2014/0218233 A1Michael PearsonPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Acute PancreatitisDocument46 pagesAcute PancreatitisLew NianPas encore d'évaluation
- Nimesulide ODTDocument10 pagesNimesulide ODTMihai SebastianPas encore d'évaluation
- Related Coverage: Stigma Experienced by Children and Adolescents With Obesity Use Neutral WordsDocument5 pagesRelated Coverage: Stigma Experienced by Children and Adolescents With Obesity Use Neutral WordsSlaghard AnerajPas encore d'évaluation
- Additive Substances in Doping: State University of JakartaDocument11 pagesAdditive Substances in Doping: State University of Jakartatiaranbl2023Pas encore d'évaluation
- Amenorrhea - Algorithm & DifferentialsDocument9 pagesAmenorrhea - Algorithm & DifferentialsItharshan IndreswaranPas encore d'évaluation
- HNS 107Document95 pagesHNS 107claudette1882Pas encore d'évaluation
- Practical Radiotherapy Planning, Fourth EditionDocument477 pagesPractical Radiotherapy Planning, Fourth EditionMehtap Coskun100% (12)
- Management of Patients With Gastric and Duodenal DisordersDocument47 pagesManagement of Patients With Gastric and Duodenal DisordersJor GarciaPas encore d'évaluation
- Immune Response in The Skin of AgingDocument7 pagesImmune Response in The Skin of AgingInmunoBlogPas encore d'évaluation
- Bladder Infection Nursing CareDocument8 pagesBladder Infection Nursing CareJoseph Anthony Benitez VerzosaPas encore d'évaluation
- EE-water Treatment-5Document13 pagesEE-water Treatment-5henil dhananiPas encore d'évaluation
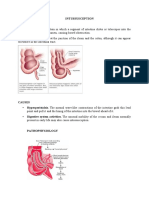
- INTUSSUSCEPTIONDocument3 pagesINTUSSUSCEPTIONS GPas encore d'évaluation
- Morris Fishbein - How the AMA Enemy Fought American Health InnovationsDocument9 pagesMorris Fishbein - How the AMA Enemy Fought American Health InnovationsuncoveringconsciousPas encore d'évaluation
- Gabapantin and PregabalinDocument23 pagesGabapantin and PregabalinayushiPas encore d'évaluation
- Yakima Co (WA) Protocols 2010Document117 pagesYakima Co (WA) Protocols 2010Anthony James-Hartwell100% (1)
- Access Medicine - TableDocument2 pagesAccess Medicine - Tablecode-24Pas encore d'évaluation
- Betamethasone (Drug Study)Document4 pagesBetamethasone (Drug Study)Franz.thenurse6888100% (1)
- Catalogo FS 07-09-2023Document10 pagesCatalogo FS 07-09-2023narciso javier Avila BejaranoPas encore d'évaluation
- Music Therapy Research PaperDocument7 pagesMusic Therapy Research Paperapi-300510538Pas encore d'évaluation
- Why Dr. Shelton Decided To Discontinue EnemasDocument3 pagesWhy Dr. Shelton Decided To Discontinue EnemasJonas Sunshine Callewaert100% (2)
- Optimal Protein Intake and The Recommended Daily Allowance (RDA)Document51 pagesOptimal Protein Intake and The Recommended Daily Allowance (RDA)BeneVia by HealthSpan InstitutePas encore d'évaluation
- Being Afraid: Everyday WorriesDocument10 pagesBeing Afraid: Everyday Worriesdina171279Pas encore d'évaluation
- Osmotic DiureticsDocument30 pagesOsmotic Diureticsrhimineecat71100% (1)
- Ai 4 PlumpynutDocument7 pagesAi 4 PlumpynutdedrascPas encore d'évaluation
- Legalization of Medical Marijuana in The Philippines H.R. 4477Document2 pagesLegalization of Medical Marijuana in The Philippines H.R. 4477Joyce Ann Mauricio50% (2)
- Hazelnuts - 7 Benefits of These Heart-Healthy, Brain-Boosting Nuts - DRDocument18 pagesHazelnuts - 7 Benefits of These Heart-Healthy, Brain-Boosting Nuts - DRRajib DasPas encore d'évaluation
- HmisDocument14 pagesHmis166873100% (1)
- Homeopathic Remedy Price List 2009-10Document1 pageHomeopathic Remedy Price List 2009-10yvranprasad100% (1)
- Art Therapy With Young Survivors of Sexual Abuse - Lost For WordsDocument215 pagesArt Therapy With Young Survivors of Sexual Abuse - Lost For WordskhalsomaliPas encore d'évaluation
- Hutzell Workbook 2009 PDFDocument48 pagesHutzell Workbook 2009 PDFPedro Almeida100% (2)