Vous aimerez peut-être aussi
- Social Isolation Selectively Reduces Hip PDFDocument3 pagesSocial Isolation Selectively Reduces Hip PDFJean Pierre Chastre LuzaPas encore d'évaluation
- Ramos Et Al 2007 DefinitivoDocument9 pagesRamos Et Al 2007 Definitivolucasmonje1978Pas encore d'évaluation
- tmp1B62 TMPDocument10 pagestmp1B62 TMPFrontiersPas encore d'évaluation
- Rebecca Bork Final MagellanDocument5 pagesRebecca Bork Final Magellanapi-736917344Pas encore d'évaluation
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenPas encore d'évaluation
- An Evaluation of Neuroplasticity and Behavior After Deep Brain Stimulation of The Nucleus Accumbens in An Animal Model of DepressionDocument10 pagesAn Evaluation of Neuroplasticity and Behavior After Deep Brain Stimulation of The Nucleus Accumbens in An Animal Model of DepressionFrontiersPas encore d'évaluation
- Epigenetic Regulation in the Nervous System: Basic Mechanisms and Clinical ImpactD'EverandEpigenetic Regulation in the Nervous System: Basic Mechanisms and Clinical ImpactPas encore d'évaluation
- Redlich 2020Document8 pagesRedlich 2020MARIA MONTSERRAT SOMOZA MONCADAPas encore d'évaluation
- Effects of Escitalopram On Synaptic Density in The HealthyDocument8 pagesEffects of Escitalopram On Synaptic Density in The Healthyarmando duvalPas encore d'évaluation
- Molecular and Cellular NeuroscienceDocument8 pagesMolecular and Cellular NeuroscienceDefi DestyawenyPas encore d'évaluation
- DurrmeyerDocument11 pagesDurrmeyermustafasacarPas encore d'évaluation
- The Brain-Derived Neurotrophic FactorDocument7 pagesThe Brain-Derived Neurotrophic FactorAlfredo E. CoronaPas encore d'évaluation
- Apha 13440Document17 pagesApha 13440javillusPas encore d'évaluation
- Aberrant Development of Intrinsic Brain Activity in A RatDocument9 pagesAberrant Development of Intrinsic Brain Activity in A RatFelipe MendesPas encore d'évaluation
- Neurplast en El Estres y ATDduman1999 PDFDocument11 pagesNeurplast en El Estres y ATDduman1999 PDFmarielaPas encore d'évaluation
- Molteni Exercise Increase Axonal RegenerationDocument6 pagesMolteni Exercise Increase Axonal RegenerationabdulPas encore d'évaluation
- The Influence of Levetiracetam in Cognitive Performance in Healthy Individuals: Neuropsychological, Behavioral and Electrophysiological ApproachDocument11 pagesThe Influence of Levetiracetam in Cognitive Performance in Healthy Individuals: Neuropsychological, Behavioral and Electrophysiological ApproachDHIVYAPas encore d'évaluation
- 1) Yan (2014)Document8 pages1) Yan (2014)Êndel AlvesPas encore d'évaluation
- Cao 2019Document11 pagesCao 2019Gustavo DídimoPas encore d'évaluation
- Pi 11 297Document10 pagesPi 11 297Sufia NaseemPas encore d'évaluation
- Dietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceDocument9 pagesDietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceJean Pierre Chastre LuzaPas encore d'évaluation
- Avanços em Psiquiatria Molecular - Março de 2023 em PDFDocument4 pagesAvanços em Psiquiatria Molecular - Março de 2023 em PDFfabriciobiblosPas encore d'évaluation
- Pnas 1305814110Document6 pagesPnas 1305814110Daniel Nuno Vancetto BorgesPas encore d'évaluation
- Palmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionDocument9 pagesPalmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionEdwing Arciniegas CarreñoPas encore d'évaluation
- The Effects of Repeated Restraint Stress On The Synaptic Plasticity in The Inner Molecular Layer of Mouse Dentate GyrusDocument13 pagesThe Effects of Repeated Restraint Stress On The Synaptic Plasticity in The Inner Molecular Layer of Mouse Dentate Gyrussonjeonggyu87Pas encore d'évaluation
- Nutritional FactorsDocument8 pagesNutritional FactorsMagnusAngeltveitKjølenPas encore d'évaluation
- Biomarkers in Overactive Bladder: Mini ReviewDocument5 pagesBiomarkers in Overactive Bladder: Mini Reviewmardha tillahPas encore d'évaluation
- LingDS 2019Document1 pageLingDS 2019NoaPas encore d'évaluation
- Role of MLC901 in Increasing Neurogenesis in Rats With Traumatic Brain InjuryDocument5 pagesRole of MLC901 in Increasing Neurogenesis in Rats With Traumatic Brain InjuryInggitaDarmawanPas encore d'évaluation
- Mehu258 U7 T25 Articulo4 PsicoterapiaDocument9 pagesMehu258 U7 T25 Articulo4 PsicoterapiaCarlos Enrique Quispe PalaciosPas encore d'évaluation
- PV Interneuron Alterations in Stress RelDocument98 pagesPV Interneuron Alterations in Stress RelSea SaltPas encore d'évaluation
- Temporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary RunningDocument8 pagesTemporal Changes of Post Synaptic Signaling Molecules, Post Synaptic Density-95 and Neuronal Nitric Oxide Synthase, in The Inner Molecular Layer of The Mouse Dentate Gyrus During Voluntary Runningsonjeonggyu87Pas encore d'évaluation
- Symposium 2023 AbstractsDocument21 pagesSymposium 2023 Abstractsmail2umamaheswari15Pas encore d'évaluation
- Beneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedDocument12 pagesBeneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedMARIA PAULA WILCHES ESCOBARPas encore d'évaluation
- 10 1016@j Euroneuro 2018 11 841Document2 pages10 1016@j Euroneuro 2018 11 841gabrielledireitoPas encore d'évaluation
- Ramirez 2015Document17 pagesRamirez 2015jarecot100% (1)
- Methylphenidate Evidence of Different Action MechanismDocument22 pagesMethylphenidate Evidence of Different Action Mechanismrafasp.arqtPas encore d'évaluation
- The Molecular Pathology of Schizophrenia: An Overview of Existing Knowledge and New Directions For Future ResearchDocument22 pagesThe Molecular Pathology of Schizophrenia: An Overview of Existing Knowledge and New Directions For Future ResearchJorge SalazarPas encore d'évaluation
- Effects of Short-Term Administration of Haloperidol On The Background of Impairments To Brain Metabolism and Neuron ActivityDocument6 pagesEffects of Short-Term Administration of Haloperidol On The Background of Impairments To Brain Metabolism and Neuron ActivitynurlithaspdnPas encore d'évaluation
- Journal of Pharmaceutical and Biomedical AnalysisDocument8 pagesJournal of Pharmaceutical and Biomedical AnalysisUday BaruahPas encore d'évaluation
- Brainsci 13 00108Document12 pagesBrainsci 13 00108Ignacio BarbieriPas encore d'évaluation
- Xiao Yao San Against Corticosterone-Induced StressDocument15 pagesXiao Yao San Against Corticosterone-Induced StressJamila IthaiaPas encore d'évaluation
- tmp490 TMPDocument11 pagestmp490 TMPFrontiersPas encore d'évaluation
- p75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor CellsDocument10 pagesp75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor Cellsmarej312Pas encore d'évaluation
- Electroconvulsive Therapy Induces Neurogenesis in Frontal Rat Brain AreasDocument5 pagesElectroconvulsive Therapy Induces Neurogenesis in Frontal Rat Brain AreasNebel GesangPas encore d'évaluation
- Norepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaDocument12 pagesNorepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaMariann DuzzPas encore d'évaluation
- Introduction To Neural Plasticity MechanismDocument9 pagesIntroduction To Neural Plasticity MechanismAlain CésarPas encore d'évaluation
- Maness EB, Et Al - Behav Brain Res. 2023 Jul 26 450 114497.Document11 pagesManess EB, Et Al - Behav Brain Res. 2023 Jul 26 450 114497.刘林玉Pas encore d'évaluation
- 2010 BP 扣带回相关的皮层结构刺激Document8 pages2010 BP 扣带回相关的皮层结构刺激XIN LVPas encore d'évaluation
- 1 s2.0 S0005796721001261 Main PDFDocument8 pages1 s2.0 S0005796721001261 Main PDFalbertoPas encore d'évaluation
- Gastrodin Promotes The Secretion of Brain-Derived Neurotrophic Factor in The Injured Spinal CordDocument17 pagesGastrodin Promotes The Secretion of Brain-Derived Neurotrophic Factor in The Injured Spinal CordAdonai AlvesPas encore d'évaluation
- Artículo NitrosynapsinDocument12 pagesArtículo NitrosynapsinViviane BocalonPas encore d'évaluation
- Clinical Efficacy and Potential Mechanisms of NeurofeedbackDocument11 pagesClinical Efficacy and Potential Mechanisms of NeurofeedbackFelix NogueiraPas encore d'évaluation
- Estrada FS Et Al Glial Activation in Pilocarpine 2012Document6 pagesEstrada FS Et Al Glial Activation in Pilocarpine 2012documentin2010Pas encore d'évaluation
- 20 - 1-S2.0-S1074742720300885-MainDocument14 pages20 - 1-S2.0-S1074742720300885-MainAna Maria RaymundiPas encore d'évaluation
- Reversing A Model of Parkinson's Disease With in Situ Converted Nigral NeuronsDocument49 pagesReversing A Model of Parkinson's Disease With in Situ Converted Nigral NeuronsPablo OrdazPas encore d'évaluation
- Genetic Reprogramming of Somatic Cells Into Neuroblasts Through - 2019 - MedicaDocument7 pagesGenetic Reprogramming of Somatic Cells Into Neuroblasts Through - 2019 - MedicaDivine Mercy Apostolate UKPas encore d'évaluation
- Article Ejbps Volume 6 August Issue 8 1564793897Document8 pagesArticle Ejbps Volume 6 August Issue 8 1564793897Rashid IqbalPas encore d'évaluation
- Personality and Individual Differences: Sharon NivDocument11 pagesPersonality and Individual Differences: Sharon NivAndrea ReisPas encore d'évaluation
- 1 s2.0 S0896627314001718 MainDocument18 pages1 s2.0 S0896627314001718 MainAhmet KayalıPas encore d'évaluation
- Insect Bites222Document5 pagesInsect Bites222Vania EssiandaPas encore d'évaluation
- Stress and Resilence With AgingDocument5 pagesStress and Resilence With AgingArouetPas encore d'évaluation
- Review Article: Rheumatoid Factors: Clinical ApplicationsDocument9 pagesReview Article: Rheumatoid Factors: Clinical ApplicationsOcef GrahastamaPas encore d'évaluation
- Regional AnesthesiaDocument24 pagesRegional AnesthesiaArouetPas encore d'évaluation
- General Studies ChecksheetDocument1 pageGeneral Studies ChecksheetArouetPas encore d'évaluation
- Journal of A Ffective Disorders: Julia C. Poole, Keith S. Dobson, Dennis PuschDocument9 pagesJournal of A Ffective Disorders: Julia C. Poole, Keith S. Dobson, Dennis PuschArouetPas encore d'évaluation
- 10.1007@s00401 016 1645 y PDFDocument23 pages10.1007@s00401 016 1645 y PDFArouetPas encore d'évaluation
- Blockposter 211424Document13 pagesBlockposter 211424ArouetPas encore d'évaluation
- 10 1016@j Molbiopara 2015 11 004 PDFDocument10 pages10 1016@j Molbiopara 2015 11 004 PDFArouetPas encore d'évaluation
- Plateefficiency Correlation in GAS P !) : Distllling AND AbsorbersDocument13 pagesPlateefficiency Correlation in GAS P !) : Distllling AND AbsorbersArouetPas encore d'évaluation
- Philosophy of MedicineDocument13 pagesPhilosophy of MedicineArouetPas encore d'évaluation
- The Oldham Era T: TH He e Ol Old DH Haam M E ErraaDocument28 pagesThe Oldham Era T: TH He e Ol Old DH Haam M E ErraaArouetPas encore d'évaluation
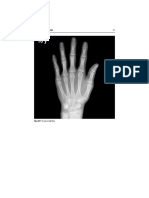
- Radiology of HandDocument1 pageRadiology of HandArouetPas encore d'évaluation
- Radiology of The HandDocument1 pageRadiology of The HandArouetPas encore d'évaluation
- 10.1007@s00401 016 1645 yDocument23 pages10.1007@s00401 016 1645 yArouetPas encore d'évaluation
- Radiology of The HandDocument1 pageRadiology of The HandArouetPas encore d'évaluation
- Radiology of HandDocument1 pageRadiology of HandArouetPas encore d'évaluation
- Radiology of The HandDocument1 pageRadiology of The HandArouetPas encore d'évaluation
- Radiology of HandDocument1 pageRadiology of HandArouetPas encore d'évaluation
- Radiology of The HandDocument1 pageRadiology of The HandArouetPas encore d'évaluation
- DenverIItest Form S PDFDocument2 pagesDenverIItest Form S PDFGaby XónPas encore d'évaluation
- Mp2016107a PDFDocument6 pagesMp2016107a PDFArouetPas encore d'évaluation
- Smith 2016Document3 pagesSmith 2016ArouetPas encore d'évaluation
- 1 STI-Estimates-Fact-Sheet-Feb-2013 PDFDocument4 pages1 STI-Estimates-Fact-Sheet-Feb-2013 PDFArouetPas encore d'évaluation
- Advanced Photography Camera Skills PDFDocument68 pagesAdvanced Photography Camera Skills PDFArouetPas encore d'évaluation
- Buono Core 2015Document2 pagesBuono Core 2015ArouetPas encore d'évaluation
- Substance P Equals Pain Substance?: News and ViewsDocument2 pagesSubstance P Equals Pain Substance?: News and ViewsArouetPas encore d'évaluation
- Roy 2016Document18 pagesRoy 2016ArouetPas encore d'évaluation
- Trikafta Combination Therapy For Cystic FibrosisDocument6 pagesTrikafta Combination Therapy For Cystic Fibrosissamarah arjumandPas encore d'évaluation
- Gene MappingDocument14 pagesGene MappingPROFESSORPas encore d'évaluation
- Fbioe 08 599674Document16 pagesFbioe 08 599674Tanishka AggarwalPas encore d'évaluation
- All Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-MullaDocument45 pagesAll Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-Mullariskhapangestika100% (1)
- Mazon - Prelims Histology Week 1Document2 pagesMazon - Prelims Histology Week 1Mazon, Dinah Melisse P.Pas encore d'évaluation
- 1 s2.0 S2352873716300166 Main PDFDocument8 pages1 s2.0 S2352873716300166 Main PDFChalisa ThamdamrongPas encore d'évaluation
- HSB Grade 10 Six WeeklyDocument5 pagesHSB Grade 10 Six WeeklyNicketa AndersonPas encore d'évaluation
- Introduction To Skin: Dr. Somaya Taha SaleemDocument17 pagesIntroduction To Skin: Dr. Somaya Taha Saleemragnarok meroPas encore d'évaluation
- GMX - BD Rhapsody Single Cell Analysis System - BR - ENDocument8 pagesGMX - BD Rhapsody Single Cell Analysis System - BR - ENMatthew GatesPas encore d'évaluation
- Biology ReviewDocument197 pagesBiology ReviewEden Lucas Ginez PitpitanPas encore d'évaluation
- Benefits of Branched Chain Amino AcidsDocument2 pagesBenefits of Branched Chain Amino AcidsparidhiPas encore d'évaluation
- Papaya Case StudyDocument4 pagesPapaya Case StudyGlowstarPas encore d'évaluation
- Platelet Physiology: Thomas Gremmel, MD Andrew L. Frelinger Iii, PHD Alan D. Michelson, MDDocument14 pagesPlatelet Physiology: Thomas Gremmel, MD Andrew L. Frelinger Iii, PHD Alan D. Michelson, MDRochnald PigaiPas encore d'évaluation
- Anti Viral DrugsDocument70 pagesAnti Viral DrugsDereje DZ100% (2)
- MalariaDocument15 pagesMalariaEssa KhanPas encore d'évaluation
- Mitochondrial DisorderDocument3 pagesMitochondrial DisorderRoxie May Theresse AbagatnanPas encore d'évaluation
- Chapter 24 DNA Viruses That Infect HumansDocument64 pagesChapter 24 DNA Viruses That Infect HumansJJ AlmagroPas encore d'évaluation
- ANALISIS HUBUNGAN KEKERABATAN TUMBUHAN BUJANG KALAM Stachytarpheta YANG DITEMUKAN DI KOTA PADANGDocument4 pagesANALISIS HUBUNGAN KEKERABATAN TUMBUHAN BUJANG KALAM Stachytarpheta YANG DITEMUKAN DI KOTA PADANGJackson SaputraPas encore d'évaluation
- I. Viruses: HIV (Human Immunodeficiency Virus)Document48 pagesI. Viruses: HIV (Human Immunodeficiency Virus)blueblemishPas encore d'évaluation
- 27 MycologyDocument55 pages27 MycologyStephen Jao Ayala UjanoPas encore d'évaluation
- Energy System: Physical Education 2Document24 pagesEnergy System: Physical Education 2Marieee Bearrr100% (1)
- Biochemistry of Liver: Alice SkoumalováDocument37 pagesBiochemistry of Liver: Alice Skoumalováandreas kevinPas encore d'évaluation
- Program Outcomes (PO's), Program Specific Outcomes (PSO's) and Course Outcomes (CO's)Document35 pagesProgram Outcomes (PO's), Program Specific Outcomes (PSO's) and Course Outcomes (CO's)Mukesh TiwariPas encore d'évaluation
- 11th STD Bio Botany TxtbookDocument336 pages11th STD Bio Botany Txtbookdravid014Pas encore d'évaluation
- Cell TheoryDocument13 pagesCell TheoryElisha TanPas encore d'évaluation
- Algae NotesDocument20 pagesAlgae NotesDHARMARAj100% (2)
- Maranatha Christian Academy SECOND Quarter Examination Science 6 I. MULTIPLE CHOICE: Choose The Letter of The Correct AnswerDocument3 pagesMaranatha Christian Academy SECOND Quarter Examination Science 6 I. MULTIPLE CHOICE: Choose The Letter of The Correct AnswerAna GamboaPas encore d'évaluation
- AN Effects of Low A260A230 012018 Ratios LRDocument6 pagesAN Effects of Low A260A230 012018 Ratios LRAndreas HaryonoPas encore d'évaluation
- Drug Discovery: Finding A Lead: An Introduction To Medicinal Chemistry 3/eDocument26 pagesDrug Discovery: Finding A Lead: An Introduction To Medicinal Chemistry 3/eMima AzrahPas encore d'évaluation