Vous aimerez peut-être aussi
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Checklist Code ReviewDocument2 pagesChecklist Code ReviewTrang Đỗ Thu100% (1)
- Mathematical Method For Physicists Ch. 1 & 2 Selected Solutions Webber and ArfkenDocument7 pagesMathematical Method For Physicists Ch. 1 & 2 Selected Solutions Webber and ArfkenJosh Brewer100% (3)
- BTL Marketing CompanyDocument30 pagesBTL Marketing Companypradip_kumarPas encore d'évaluation
- Neurodegeneration in Diabetic RetinopathyDocument11 pagesNeurodegeneration in Diabetic RetinopathyKahfi RakhmadianPas encore d'évaluation
- Multiple Trauma TKVDocument21 pagesMultiple Trauma TKVKahfi RakhmadianPas encore d'évaluation
- Wednesday, May 18 2022: Duty ReportDocument60 pagesWednesday, May 18 2022: Duty ReportKahfi RakhmadianPas encore d'évaluation
- Omega-3 Fatty Acids For Breast Cancer PreventionDocument11 pagesOmega-3 Fatty Acids For Breast Cancer PreventionKahfi RakhmadianPas encore d'évaluation
- Filename 0 Effect of Polyunsaturated Omega-3 and Omega-6 Fatty Acids oDocument20 pagesFilename 0 Effect of Polyunsaturated Omega-3 and Omega-6 Fatty Acids oKahfi RakhmadianPas encore d'évaluation
- Helping Women To Good Health Breast CancerDocument5 pagesHelping Women To Good Health Breast CancerKahfi RakhmadianPas encore d'évaluation
- Distribution of Prolactin Receptors Suggests AnDocument15 pagesDistribution of Prolactin Receptors Suggests AnKahfi RakhmadianPas encore d'évaluation
- 05 PetricevicDocument8 pages05 PetricevicKahfi RakhmadianPas encore d'évaluation
- Gain-Of-function Prolactin Receptor Variants Are NDocument11 pagesGain-Of-function Prolactin Receptor Variants Are NKahfi RakhmadianPas encore d'évaluation
- Plasma Prolactin and Breast Cancer Riska Meta - AnalysisDocument7 pagesPlasma Prolactin and Breast Cancer Riska Meta - AnalysisKahfi RakhmadianPas encore d'évaluation
- Prolactin Receptor Breast Cancer PremenopausalDocument9 pagesProlactin Receptor Breast Cancer PremenopausalKahfi RakhmadianPas encore d'évaluation
- S L Dixon Fluid Mechanics and Thermodynamics of TurbomachineryDocument4 pagesS L Dixon Fluid Mechanics and Thermodynamics of Turbomachinerykuma alemayehuPas encore d'évaluation
- 11 Stem P - Group 2 - CPT First GradingDocument7 pages11 Stem P - Group 2 - CPT First GradingZwen Zyronne Norico LumiwesPas encore d'évaluation
- Veris Case StudyDocument2 pagesVeris Case StudyPankaj GargPas encore d'évaluation
- Consumer PresentationDocument30 pagesConsumer PresentationShafiqur Rahman KhanPas encore d'évaluation
- AppendixA LaplaceDocument12 pagesAppendixA LaplaceSunny SunPas encore d'évaluation
- CFA L1 Ethics Questions and AnswersDocument94 pagesCFA L1 Ethics Questions and AnswersMaulik PatelPas encore d'évaluation
- OD - SAP Connector UtilityDocument22 pagesOD - SAP Connector UtilityShivani SharmaPas encore d'évaluation
- BMW Engine Wiring 2 OxDocument4 pagesBMW Engine Wiring 2 OxTomáš TdcPas encore d'évaluation
- Festival Implementation PlanDocument5 pagesFestival Implementation Planapi-318058589Pas encore d'évaluation
- Annexure I Project Details DateDocument4 pagesAnnexure I Project Details DateAshish SinghaniaPas encore d'évaluation
- 1 s2.0 S0955221920305689 MainDocument19 pages1 s2.0 S0955221920305689 MainJoaoPas encore d'évaluation
- APPSC GR I Initial Key Paper IIDocument52 pagesAPPSC GR I Initial Key Paper IIdarimaduguPas encore d'évaluation
- English Literature Coursework Aqa GcseDocument6 pagesEnglish Literature Coursework Aqa Gcsef5d17e05100% (2)
- Brain Alchemy Masterclass PsychotacticsDocument87 pagesBrain Alchemy Masterclass Psychotacticskscmain83% (6)
- Strategy Guide To Twilight Imperium Third EditionDocument74 pagesStrategy Guide To Twilight Imperium Third Editioninquartata100% (1)
- 2062 TSSR Site Sharing - Rev02Document44 pages2062 TSSR Site Sharing - Rev02Rio DefragPas encore d'évaluation
- RISO RZ User GuideDocument112 pagesRISO RZ User GuideJojo AritallaPas encore d'évaluation
- Case Study StarbucksDocument2 pagesCase Study StarbucksSonal Agarwal100% (2)
- CAKUTDocument50 pagesCAKUTsantosh subediPas encore d'évaluation
- 039-Itp For Embedded Plate PDFDocument7 pages039-Itp For Embedded Plate PDFKöksal PatanPas encore d'évaluation
- Stadium and Club Tours - Senior Tour GuideDocument4 pagesStadium and Club Tours - Senior Tour GuidebizmbuuPas encore d'évaluation
- 10.ULABs Presentation Camiguin FinalDocument55 pages10.ULABs Presentation Camiguin FinalKaren Feyt MallariPas encore d'évaluation
- A Modified Linear Programming Method For Distribution System ReconfigurationDocument6 pagesA Modified Linear Programming Method For Distribution System Reconfigurationapi-3697505Pas encore d'évaluation
- Biology Concepts and Applications 9th Edition Starr Solutions ManualDocument9 pagesBiology Concepts and Applications 9th Edition Starr Solutions Manualscarletwilliamnfz100% (31)
- Preparation of Kevlar-49 Fabric/E-Glass Fabric/Epoxy Composite Materials and Characterization of Their Mechanical PropertiesDocument10 pagesPreparation of Kevlar-49 Fabric/E-Glass Fabric/Epoxy Composite Materials and Characterization of Their Mechanical PropertiesIndhu ElangoPas encore d'évaluation
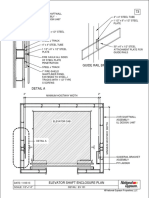
- Guide Rail Bracket AssemblyDocument1 pageGuide Rail Bracket AssemblyPrasanth VarrierPas encore d'évaluation
- 05271/MFP YPR SPL Sleeper Class (SL)Document2 pages05271/MFP YPR SPL Sleeper Class (SL)Rdx BoePas encore d'évaluation