Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- All About WomenDocument96 pagesAll About Womenex360100% (1)
- Autopsy of Baby WyllowDocument10 pagesAutopsy of Baby WyllowKOLD News 13100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- Ashringa: 5 Horses of Thunder Totems & SpiritsDocument16 pagesAshringa: 5 Horses of Thunder Totems & SpiritsLauraHenson100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- SportDog Brand Basic TrainingDocument19 pagesSportDog Brand Basic TrainingkwrdkcPas encore d'évaluation
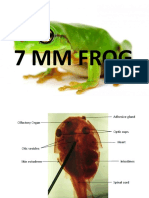
- 7 MM FrogDocument28 pages7 MM FrogNiki Reroll04Pas encore d'évaluation
- Allan Savory - Holistic Management OverviewDocument90 pagesAllan Savory - Holistic Management OverviewLady Galadriel100% (2)
- PASTURE MANAGEMENT - Final PDFDocument101 pagesPASTURE MANAGEMENT - Final PDFDexter Andrew Lyken100% (1)
- Pastest 2016-2017-AKP - Diabetes and EndocrineDocument62 pagesPastest 2016-2017-AKP - Diabetes and EndocrineHisham Elkilaney100% (1)
- Anal FissureDocument16 pagesAnal FissureJehanzeb Zeb100% (1)
- My Daddy S Best FriendDocument32 pagesMy Daddy S Best Friendhyunheejo98Pas encore d'évaluation
- What Is Hypotonic SolutionDocument5 pagesWhat Is Hypotonic SolutionC Six Nor AzwaniPas encore d'évaluation
- Recurrent Vulvovaginal Candidiasis: Evidence-Based Pharmacy PracticeDocument4 pagesRecurrent Vulvovaginal Candidiasis: Evidence-Based Pharmacy PracticeRiddhi JainPas encore d'évaluation
- The Voyages of Sinbad The Sailor. Answer KeyDocument1 pageThe Voyages of Sinbad The Sailor. Answer KeyOleg MuhendisPas encore d'évaluation
- PM Artifacts WEBDocument20 pagesPM Artifacts WEBSalman MemonPas encore d'évaluation
- Hygienic Practices in Slaughter HouseDocument40 pagesHygienic Practices in Slaughter Housezubair valiPas encore d'évaluation
- Ansci 1105 ReviewerDocument13 pagesAnsci 1105 ReviewerCj M SapadPas encore d'évaluation
- Long Quiz 1 BiologyDocument2 pagesLong Quiz 1 BiologyNonita Caracas CubiloPas encore d'évaluation
- MICRO Lecture 1 Introduction To Microbiology and Parasitology 1 PDFDocument29 pagesMICRO Lecture 1 Introduction To Microbiology and Parasitology 1 PDFJireh AcabalPas encore d'évaluation
- Whats The SettingDocument2 pagesWhats The Settingapi-2428046300% (2)
- Soal TOEFLDocument2 pagesSoal TOEFLArraya Eritha BarcelonaPas encore d'évaluation
- Story Comm #150: "A Show For Charlotte": (Art by Mastergodai)Document6 pagesStory Comm #150: "A Show For Charlotte": (Art by Mastergodai)Szymon SłomskiPas encore d'évaluation
- Now Begin Work On The QuestionsDocument4 pagesNow Begin Work On The QuestionsICaa Dwiiy AnndiechtyaPas encore d'évaluation
- Assessment Goal 7Document24 pagesAssessment Goal 7api-333352303Pas encore d'évaluation
- Richard Ryder: All Beings That Feel Pain Deserve Human Rights. The GuardianDocument3 pagesRichard Ryder: All Beings That Feel Pain Deserve Human Rights. The GuardianpilisantaPas encore d'évaluation
- Optimal Sizing of Counterflow Cooler For PelletsDocument72 pagesOptimal Sizing of Counterflow Cooler For PelletsChristian MavarezPas encore d'évaluation
- Dual CementDocument33 pagesDual CementStephani CastilloPas encore d'évaluation
- Standards For Blood Banks and Transfusion Services Standards For Blood Banks and Transfusion ServicesDocument5 pagesStandards For Blood Banks and Transfusion Services Standards For Blood Banks and Transfusion Serviceslaexm95Pas encore d'évaluation
- Behavioral and Motivational Mechanisms of The BrainDocument58 pagesBehavioral and Motivational Mechanisms of The BrainLarry TongPas encore d'évaluation
- Pengaruh Umur Induk Dan Lama Penyimpanan Telur Terhadap Bobot Telur, Daya Tetas Dan Bobot Tetas Ayam Kedu Jengger MerahDocument4 pagesPengaruh Umur Induk Dan Lama Penyimpanan Telur Terhadap Bobot Telur, Daya Tetas Dan Bobot Tetas Ayam Kedu Jengger MerahKKN-T Gorontalo 2020Pas encore d'évaluation
- Dukan Diet Food ListDocument2 pagesDukan Diet Food ListprogramgrabberPas encore d'évaluation