Vous aimerez peut-être aussi
- Chemical Signal: Hormone 1: Biosignaling (WEEK 2)Document27 pagesChemical Signal: Hormone 1: Biosignaling (WEEK 2)Fenny Aulia SugianaPas encore d'évaluation
- Conversion RPM G CentrifugaDocument1 pageConversion RPM G CentrifugaEsaú E RodriguezPas encore d'évaluation
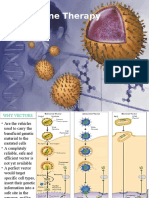
- Gene Therapy For Hemophilia: Marinee K. L. Chuah Desire Collen Thierry VandendriesscheDocument18 pagesGene Therapy For Hemophilia: Marinee K. L. Chuah Desire Collen Thierry VandendriesscheFenny Aulia SugianaPas encore d'évaluation
- Vectors in Gene TherapyDocument37 pagesVectors in Gene TherapyFenny Aulia SugianaPas encore d'évaluation
- Articol Exosomi ImpDocument11 pagesArticol Exosomi Impanon_579968442Pas encore d'évaluation
- IMVICDocument20 pagesIMVICFenny Aulia Sugiana100% (1)
- B2 SEC BiophysJ Oct 03Document5 pagesB2 SEC BiophysJ Oct 03Fenny Aulia SugianaPas encore d'évaluation
- Guru Nanak Institute of Pharmacy, Ibrahimpatnam, Hyderabad-501506, Dist-Ranga Reddy, Andhra PradeshDocument9 pagesGuru Nanak Institute of Pharmacy, Ibrahimpatnam, Hyderabad-501506, Dist-Ranga Reddy, Andhra PradeshNur Aini IktikhafsariPas encore d'évaluation
- Crcsodium Review PTS - 391-416Document27 pagesCrcsodium Review PTS - 391-416Fenny Aulia SugianaPas encore d'évaluation
- Jurnal LKM 2Document12 pagesJurnal LKM 2PIt Season IIPas encore d'évaluation
- Effect of High-Fat Diets On Body Composition, Lipid Metabolism and Insulin Sensitivity, and The Role of Exercise On These ParametersDocument8 pagesEffect of High-Fat Diets On Body Composition, Lipid Metabolism and Insulin Sensitivity, and The Role of Exercise On These ParametersFenny Aulia SugianaPas encore d'évaluation
- IMVICDocument20 pagesIMVICFenny Aulia Sugiana100% (1)
- Jbct12i2p121 PDFDocument4 pagesJbct12i2p121 PDFFenny Aulia SugianaPas encore d'évaluation
- Sample Preparation Techniques in Biochemistry With Particular Reference To HeterogeneousDocument16 pagesSample Preparation Techniques in Biochemistry With Particular Reference To HeterogeneousFenny Aulia SugianaPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Substance Use Disorders Among Youth in The Juvenile Justice SystemDocument14 pagesSubstance Use Disorders Among Youth in The Juvenile Justice SystemJames JerryPas encore d'évaluation
- Eye Pain and DischargeDocument53 pagesEye Pain and DischargeJitendra ChaudharyPas encore d'évaluation
- Ethiopia - Design Gudeline Final - OromiaDocument71 pagesEthiopia - Design Gudeline Final - OromiaEphrem Gizachew100% (15)
- Reflective Journal 1 2 and 3Document11 pagesReflective Journal 1 2 and 3api-321980896Pas encore d'évaluation
- HSE Assignment 3 FinalDocument21 pagesHSE Assignment 3 FinalLuqman HakimPas encore d'évaluation
- X120MN12Document6 pagesX120MN12mini p shendePas encore d'évaluation
- Root Caries: A Periodontal Perspective: Review ArticleDocument21 pagesRoot Caries: A Periodontal Perspective: Review ArticleIlmia AmaliaPas encore d'évaluation
- Table Saw Safety FinalDocument24 pagesTable Saw Safety Finalvolatileyasser100% (1)
- TRICARE Program Integrity 2010 ReportDocument25 pagesTRICARE Program Integrity 2010 ReportGovtfraudlawyerPas encore d'évaluation
- Manual PTCDocument64 pagesManual PTCSeba SapiainPas encore d'évaluation
- RULE 1142. Marine Tank Vessel OperationsDocument8 pagesRULE 1142. Marine Tank Vessel OperationsksalvaradoPas encore d'évaluation
- Employee Benefits India PDFDocument2 pagesEmployee Benefits India PDFshahidki31100% (1)
- Importance of An Active LifestyleDocument2 pagesImportance of An Active LifestyleDreamCatcher Insommia18Pas encore d'évaluation
- ST Regis OriginalDocument224 pagesST Regis OriginalNeeraj AgarwalPas encore d'évaluation
- Daftar PustakaDocument3 pagesDaftar PustakaTeguh firmansyahPas encore d'évaluation
- Ijc Heart & Vasculature: SciencedirectDocument8 pagesIjc Heart & Vasculature: SciencedirectFerdi YuanPas encore d'évaluation
- Amal ResumeDocument2 pagesAmal Resumeapi-242600239Pas encore d'évaluation
- 2021 A Guide To The African Human Rights System Celebrating 40 YearsDocument115 pages2021 A Guide To The African Human Rights System Celebrating 40 YearsChala Yuye KemerPas encore d'évaluation
- Pengaruh Aplikasi Pyraclostrobin Terhadap Serangan Penyebab Penyakit Bulai Pada Lima Varietas Jagung (Zea Mays)Document8 pagesPengaruh Aplikasi Pyraclostrobin Terhadap Serangan Penyebab Penyakit Bulai Pada Lima Varietas Jagung (Zea Mays)Rajif S. YahyaPas encore d'évaluation
- NDEPDocument5 pagesNDEPjoePas encore d'évaluation
- School Canteen Feeding Program 2019 2020 K 6 1Document20 pagesSchool Canteen Feeding Program 2019 2020 K 6 1MaryRoseTrinidadPas encore d'évaluation
- Essay (DUNGARAN, JOHN KLENN) (BSCE-1CDocument2 pagesEssay (DUNGARAN, JOHN KLENN) (BSCE-1CJohn KlennPas encore d'évaluation
- Checklist Auditoria SQF 2010Document21 pagesChecklist Auditoria SQF 2010cramirez48Pas encore d'évaluation
- Pathologist Job SpecificationDocument16 pagesPathologist Job SpecificationLalaluluPas encore d'évaluation
- Tinnitus Today September 1986 Vol 11, No 3Document8 pagesTinnitus Today September 1986 Vol 11, No 3American Tinnitus AssociationPas encore d'évaluation
- Clinical Manifestations and Diagnosis of Raynaud Phenomenon - UpToDateDocument29 pagesClinical Manifestations and Diagnosis of Raynaud Phenomenon - UpToDateMarcos Torchia EstebanPas encore d'évaluation
- Genflex Roofing Systems, LLC Material Safety Data SheetDocument4 pagesGenflex Roofing Systems, LLC Material Safety Data SheetgjroddyPas encore d'évaluation
- Metaparadigm in NursingDocument3 pagesMetaparadigm in NursingJenno Ray SenalPas encore d'évaluation
- Common Sports InjuryDocument71 pagesCommon Sports InjuryGlen Dizon100% (1)
- PD 1067 Water Code of The Philippines and Clean Water ActDocument44 pagesPD 1067 Water Code of The Philippines and Clean Water ActNovy Jamboy Ragmac100% (2)