Vous aimerez peut-être aussi
- Biología C.M Seminario - Informe S3.Grupo 05Document7 pagesBiología C.M Seminario - Informe S3.Grupo 05Edwin GonzalesPas encore d'évaluation
- Actividad 5 - Lo Que Podemos ControlarDocument3 pagesActividad 5 - Lo Que Podemos ControlarPaula Lizeth Ramirez SanchezPas encore d'évaluation
- 05 Citoplasma - CitoesqueletoDocument19 pages05 Citoplasma - CitoesqueletoAlfredo Sam Alvaro HerreraPas encore d'évaluation
- Estructura y Funcion de Celulas Procariotas y EucariotasDocument6 pagesEstructura y Funcion de Celulas Procariotas y EucariotasJose Gonzales100% (1)
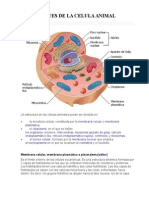
- Célula AnimalDocument17 pagesCélula AnimalJUDITH ALONDRA TORRES HERNANDEZPas encore d'évaluation
- Semana 06 ESTRUCTURA Y FUNCION CELULARDocument14 pagesSemana 06 ESTRUCTURA Y FUNCION CELULARMARKO ANTONIO RAMOS QUISPEPas encore d'évaluation
- UNIDAD 1 CitologiaDocument16 pagesUNIDAD 1 CitologiaLara AgnoluzziPas encore d'évaluation
- Seg Exa BiologiaDocument36 pagesSeg Exa BiologiaBrayan Villavicencio TovarPas encore d'évaluation
- El CitoplasmaDocument6 pagesEl Citoplasmaoscar david mercado beltranPas encore d'évaluation
- Las Entidades Fundamentales de Todo Ser Vivo Son Las CélulasDocument5 pagesLas Entidades Fundamentales de Todo Ser Vivo Son Las CélulasroximarcontrerasPas encore d'évaluation
- B2 T4 - CÉLULA EUCARIOTA - Citoplasma ApuntesDocument16 pagesB2 T4 - CÉLULA EUCARIOTA - Citoplasma ApuntesNacho Orio FernándezPas encore d'évaluation
- Citosol, Citoesqueleto Parte 1Document70 pagesCitosol, Citoesqueleto Parte 1Mildred IbañezPas encore d'évaluation
- Tipos Celulares y Célula Eucariota 22Document19 pagesTipos Celulares y Célula Eucariota 22Isis RuedaPas encore d'évaluation
- FICHA PEDAGOGICA XVI Biologia 1ro BguDocument11 pagesFICHA PEDAGOGICA XVI Biologia 1ro BguAndrea DanielaPas encore d'évaluation
- Cito EsqueletoDocument8 pagesCito EsqueletoJuan OrtízPas encore d'évaluation
- Cuidados Post MortemDocument14 pagesCuidados Post MortemAdri AdrianPas encore d'évaluation
- Elementos Citoesqueleticos y Matriz CitoplasmaticaDocument6 pagesElementos Citoesqueleticos y Matriz CitoplasmaticagiovannyPas encore d'évaluation
- La Estructura de La CélulaDocument6 pagesLa Estructura de La CélulaMaricielo Calderon MinayaPas encore d'évaluation
- Apunte La..Document11 pagesApunte La..Guillermina ArroyoPas encore d'évaluation
- Citoplasma - 2024Document13 pagesCitoplasma - 2024morena RuizPas encore d'évaluation
- 2 Trabajo - NeurocienciaDocument15 pages2 Trabajo - NeurocienciaLuciaPas encore d'évaluation
- CélulaDocument18 pagesCélulaABAD TPas encore d'évaluation
- Bio PFDocument4 pagesBio PFLizley AbigailPas encore d'évaluation
- CITOESQUELETODocument3 pagesCITOESQUELETOMarianaPas encore d'évaluation
- Célula AnimalDocument13 pagesCélula AnimalRosita SilvaPas encore d'évaluation
- Módulo 6 I La CelulaDocument77 pagesMódulo 6 I La CelulaVale Jaramillo GilPas encore d'évaluation
- Parcial BiologiaDocument24 pagesParcial BiologialorejojedaPas encore d'évaluation
- Celula AnimalDocument11 pagesCelula AnimalDania FloresPas encore d'évaluation
- La CélulaDocument14 pagesLa CélulaDeiv PerezPas encore d'évaluation
- La CélulaDocument8 pagesLa CélulaFrancis BravoPas encore d'évaluation
- Características de La Célula Animal y VegetalDocument13 pagesCaracterísticas de La Célula Animal y VegetalJehison Figueroa100% (2)
- Unidad 3 La CelulaDocument8 pagesUnidad 3 La CelulaTania LopezPas encore d'évaluation
- Organelos CelularesDocument13 pagesOrganelos CelularesEduardo Jaziel Avelino BuendiaPas encore d'évaluation
- Investigacion de La Celula HumanaDocument9 pagesInvestigacion de La Celula HumanaJuanito HernandezPas encore d'évaluation
- CELULADocument108 pagesCELULAalejandraPas encore d'évaluation
- Célula AnimalDocument5 pagesCélula AnimaljhoonnPas encore d'évaluation
- Estructura y Función Celular 1.Document22 pagesEstructura y Función Celular 1.macavecaPas encore d'évaluation
- Metabolismo VegetalDocument45 pagesMetabolismo VegetalAlejandro GuzmanPas encore d'évaluation
- Biología Clase 27.04Document18 pagesBiología Clase 27.04Camii MariqueoPas encore d'évaluation
- Documento 6Document4 pagesDocumento 6Mónica Azucena Prado GómezPas encore d'évaluation
- Organelos Membranosos y No Membranosos ScribdDocument4 pagesOrganelos Membranosos y No Membranosos ScribdJosePas encore d'évaluation
- Guia de Estudio 1biologíaDocument7 pagesGuia de Estudio 1biologíaCristina MoralesPas encore d'évaluation
- Tema 3 Estructura y Función CelularDocument8 pagesTema 3 Estructura y Función CelularJorge GarcíaPas encore d'évaluation
- Guía3 CELULADocument6 pagesGuía3 CELULAignaciaPas encore d'évaluation
- TEORÍA Citoesqueleto Plastidios, Cilios, Flagelos y CentriolosDocument82 pagesTEORÍA Citoesqueleto Plastidios, Cilios, Flagelos y CentriolosJulian HuamaniPas encore d'évaluation
- Citoplasma, Citoesqueleto y Tráfico VesicularDocument15 pagesCitoplasma, Citoesqueleto y Tráfico VesicularMel Pérez BorgesPas encore d'évaluation
- Guia Organelos CelularesDocument4 pagesGuia Organelos CelularesjonathanPas encore d'évaluation
- La CélulaDocument6 pagesLa CélulaDerek Jhonntanh Villalba SandovalPas encore d'évaluation
- Partes de La Celula AnimalDocument7 pagesPartes de La Celula AnimalDanilo Daniel López PérezPas encore d'évaluation
- Guía 4 Citoplasma y NucleoDocument13 pagesGuía 4 Citoplasma y Nucleodomenica guamanPas encore d'évaluation
- Organelos CelularesDocument12 pagesOrganelos CelularesJulio Rz100% (3)
- CitologíaDocument5 pagesCitologíaHelmer Roque GamarraPas encore d'évaluation
- Anato 1Document61 pagesAnato 1NALLELYPas encore d'évaluation
- ACFrOgAYBrPgjgUiHGJkJ304e - 6gDa6yOwClhhVSDjI9EPj7wk VL - M6iKrx5uxSJP X426peBQvUhLXWLRNmG05QdtK hgE79L6fle7fAD we7Ctoe6oCNDA3XXfnUTfcaHjERHR - ASdlW CJs PDFDocument138 pagesACFrOgAYBrPgjgUiHGJkJ304e - 6gDa6yOwClhhVSDjI9EPj7wk VL - M6iKrx5uxSJP X426peBQvUhLXWLRNmG05QdtK hgE79L6fle7fAD we7Ctoe6oCNDA3XXfnUTfcaHjERHR - ASdlW CJs PDFrosa ayanPas encore d'évaluation
- Cito PlasmaDocument8 pagesCito PlasmaWalter oliveraPas encore d'évaluation
- Histología y Embrología del Sitema NerviosoD'EverandHistología y Embrología del Sitema NerviosoÉvaluation : 4 sur 5 étoiles4/5 (4)
- Anemia Para Todos: Medicina Para TodosD'EverandAnemia Para Todos: Medicina Para TodosÉvaluation : 3 sur 5 étoiles3/5 (1)
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularD'EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularPas encore d'évaluation
- Tarea No. #1 Auditoria ComputarizadoDocument20 pagesTarea No. #1 Auditoria ComputarizadoDenian CastilloPas encore d'évaluation
- Auditoria Computarizada EjemploDocument7 pagesAuditoria Computarizada EjemploDenian CastilloPas encore d'évaluation
- Texto Paralelo Proyecto III Primera ParteDocument12 pagesTexto Paralelo Proyecto III Primera ParteDenian CastilloPas encore d'évaluation
- Rural CaratulaDocument4 pagesRural CaratulaDenian CastilloPas encore d'évaluation
- Cómo Se Calcula El ISR en GuatemalaDocument5 pagesCómo Se Calcula El ISR en GuatemalaDenian CastilloPas encore d'évaluation
- Estados FinancierosDocument39 pagesEstados FinancierosDenian CastilloPas encore d'évaluation
- Texto Paralelo Auditoria IDocument47 pagesTexto Paralelo Auditoria IDenian CastilloPas encore d'évaluation
- Cronograma de Tesis 12septiembre2016Document1 pageCronograma de Tesis 12septiembre2016Denian CastilloPas encore d'évaluation
- Texto Paralelo PlanificacionDocument50 pagesTexto Paralelo PlanificacionDenian CastilloPas encore d'évaluation
- Biologia 3 BguDocument3 pagesBiologia 3 BguElizabeth ZambranoPas encore d'évaluation
- Sistema Endocrino y CardiovascularDocument119 pagesSistema Endocrino y CardiovascularMafer Taco TanguilaPas encore d'évaluation
- Hipoaldosteronismo HiporreninemicoDocument9 pagesHipoaldosteronismo Hiporreninemico1309Pas encore d'évaluation
- Copia de Biología II - Cuaderno de Trabajo 2020Document35 pagesCopia de Biología II - Cuaderno de Trabajo 2020ROSELENA VELAZQUEZ LOPEZPas encore d'évaluation
- Parcial de Fisiio - Sistema RenalDocument4 pagesParcial de Fisiio - Sistema RenalPaulo PavoniPas encore d'évaluation
- Manual de Prácticas de Laboratorio de DisecciónDocument20 pagesManual de Prácticas de Laboratorio de DisecciónPaola VillanuevaPas encore d'évaluation
- BhujangasanaDocument7 pagesBhujangasanaIvan Calero GranadosPas encore d'évaluation
- Cuestionario Unidad VI Músculo LisoDocument3 pagesCuestionario Unidad VI Músculo LisoA.D.P.GPas encore d'évaluation
- Resumen Y Mapas - SNC-SNP-y-Piel1-equipo 4Document15 pagesResumen Y Mapas - SNC-SNP-y-Piel1-equipo 4Mauricio Caña AvalosPas encore d'évaluation
- Aparato UrinarioDocument5 pagesAparato UrinarioSofia MedinaPas encore d'évaluation
- Fisiología de La ProducciónDocument4 pagesFisiología de La ProducciónAngieCruzPas encore d'évaluation
- Médico Federico RuscittoDocument40 pagesMédico Federico RuscittoEze BasilePas encore d'évaluation
- La Verdad Sobre Los CarbohidratosDocument23 pagesLa Verdad Sobre Los Carbohidratosedgar facundoPas encore d'évaluation
- Irrigación Del CorazónDocument1 pageIrrigación Del CorazónAlexandra DuránPas encore d'évaluation
- Taller ProteínasDocument5 pagesTaller ProteínasDiego RodriguezPas encore d'évaluation
- Sentido Del OlfatoDocument6 pagesSentido Del OlfatoCALIXTOPas encore d'évaluation
- Bioenergetica y Oxidacion Biologica ResumenDocument13 pagesBioenergetica y Oxidacion Biologica ResumenPamela AquinoPas encore d'évaluation
- Cuestionario Sobre Diversidad Celular.Document7 pagesCuestionario Sobre Diversidad Celular.Andreína HigueraPas encore d'évaluation
- Preguntas GRUPO 2 CEREBRODocument10 pagesPreguntas GRUPO 2 CEREBROAllisson ZavalaPas encore d'évaluation
- 2°1° BiologiaDocument3 pages2°1° BiologiaRoque FeitoPas encore d'évaluation
- Qué y Cuáles Son Los Procesos CelularesDocument6 pagesQué y Cuáles Son Los Procesos CelularesAlexis HasePas encore d'évaluation
- Area de Hematología: Empresa: Salud Total EPS-S S.ADocument2 pagesArea de Hematología: Empresa: Salud Total EPS-S S.ACONTADOR CONTADORPas encore d'évaluation
- Tarea Del Lab. Anatomia PatologicaDocument4 pagesTarea Del Lab. Anatomia PatologicaGabriela FabianPas encore d'évaluation
- SILABO FISIOLOGÍA HUMANA 2023-II VisadoDocument32 pagesSILABO FISIOLOGÍA HUMANA 2023-II Visadoeduardo imanPas encore d'évaluation
- Taller de Enfermería Básica - Aiep - Via Parental - Enteral - FluidosDocument35 pagesTaller de Enfermería Básica - Aiep - Via Parental - Enteral - FluidosCarolina Villalobos ArayaPas encore d'évaluation
- Tarea 1 InhaloterapiaDocument59 pagesTarea 1 InhaloterapiaMarcela CendejasPas encore d'évaluation
- Cap 6Document7 pagesCap 6Natalia Espíndola BalderasPas encore d'évaluation
- Guía de FarmacologíaDocument6 pagesGuía de FarmacologíaOscar CisnerosPas encore d'évaluation
- Anatomía y FisiologíaDocument24 pagesAnatomía y FisiologíaAimeé GuerreroPas encore d'évaluation