Vous aimerez peut-être aussi
- Glutamate Ionotropic Receptors: AMPA and NMDA ReceptorsDocument3 pagesGlutamate Ionotropic Receptors: AMPA and NMDA Receptorslinnet17Pas encore d'évaluation
- NMDA Receptors in The Central Nervous SystemDocument80 pagesNMDA Receptors in The Central Nervous SystemDiana ParraPas encore d'évaluation
- Synapses: Department of Mathematical SciencesDocument7 pagesSynapses: Department of Mathematical SciencesilincaPas encore d'évaluation
- By: Prashant Kumar Singh M. Pharm (Ph. Analysis) Mangalayatan University AligarhDocument17 pagesBy: Prashant Kumar Singh M. Pharm (Ph. Analysis) Mangalayatan University AligarhPrashant SinghPas encore d'évaluation
- ReceptorsDocument9 pagesReceptorsrajenderPas encore d'évaluation
- Bahasa Inggris Stahl Ion ChanelDocument5 pagesBahasa Inggris Stahl Ion ChanelMuhammad Zul Fahmi AkbarPas encore d'évaluation
- Idk 3 Case 2 NBSDocument2 pagesIdk 3 Case 2 NBSAlit DarmayantiPas encore d'évaluation
- Medical Neuroscience Tutorial Notes: Neurotransmitter ReceptorsDocument5 pagesMedical Neuroscience Tutorial Notes: Neurotransmitter ReceptorsEdvard JamesPas encore d'évaluation
- Cousson Et Al., 1998 Two Potential...Document6 pagesCousson Et Al., 1998 Two Potential...Gian Carlo CarvalhoPas encore d'évaluation
- Gamma-Aminobutyric Acid or 4-Aminobutanoic Acid Is A Gamma-Amino Acid That Is With The Amino Substituent Located at C-4Document5 pagesGamma-Aminobutyric Acid or 4-Aminobutanoic Acid Is A Gamma-Amino Acid That Is With The Amino Substituent Located at C-4CourtneyPas encore d'évaluation
- Modulation of Channel Function by PolyaminesDocument6 pagesModulation of Channel Function by Polyaminesbrenda_bbPas encore d'évaluation
- Visual Phototransduction Is The SensoryDocument5 pagesVisual Phototransduction Is The SensoryNTA UGC-NETPas encore d'évaluation
- Metabotropic Nmda ReceptorDocument6 pagesMetabotropic Nmda ReceptorPaco Ibañez BravoPas encore d'évaluation
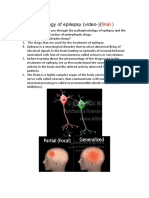
- EpilepsyDocument5 pagesEpilepsydr.pragna infoplusPas encore d'évaluation
- Types of ReceptorsDocument78 pagesTypes of ReceptorsDrPallavi DPas encore d'évaluation
- Glutamate Major Excitatory Neurotransmitter. Two Types of ReceptorsDocument2 pagesGlutamate Major Excitatory Neurotransmitter. Two Types of ReceptorsELIOPas encore d'évaluation
- Kew 2005 Ionotropic and Metabotropic Glu Rec Structure (Ábra)Document26 pagesKew 2005 Ionotropic and Metabotropic Glu Rec Structure (Ábra)zernebochPas encore d'évaluation
- Purves Neuroscience Website Questions CH 8 AnswersDocument4 pagesPurves Neuroscience Website Questions CH 8 AnswersPK3214598750% (2)
- AMPK Mechanisms of Cellular Energy Sensing N Restoration of Metabolic BalanceDocument12 pagesAMPK Mechanisms of Cellular Energy Sensing N Restoration of Metabolic BalanceOsvaldo VillarPas encore d'évaluation
- Electro-Physiology of HeartDocument7 pagesElectro-Physiology of HeartEhtiram HuseynovPas encore d'évaluation
- NTs RsDocument45 pagesNTs RsAhmed IsmailPas encore d'évaluation
- 01-02 MS Ion Channels PDFDocument30 pages01-02 MS Ion Channels PDF16_dev5038Pas encore d'évaluation
- Structural Biochemistry of Atp-Driven Dimerization and Dna-Stimulated Activation of SMC AtpasesDocument5 pagesStructural Biochemistry of Atp-Driven Dimerization and Dna-Stimulated Activation of SMC Atpasesomar ManPas encore d'évaluation
- 01-02 MS Ion Channels PDFDocument30 pages01-02 MS Ion Channels PDFSwaranjeet SinghPas encore d'évaluation
- 11 Synaptic Plasticity LTP LTDDocument4 pages11 Synaptic Plasticity LTP LTDCesia ConstanzaPas encore d'évaluation
- Purich 2010Document1 pagePurich 2010Stjepan BičanićPas encore d'évaluation
- 2 PHAR0004 9 7 ReceptorStructure NSM 2018 NotesDocument3 pages2 PHAR0004 9 7 ReceptorStructure NSM 2018 NotesArthi ArumukasamyPas encore d'évaluation
- Letter: Methanocaldococcus Jannaschii Escherichia Coli E. ColiDocument7 pagesLetter: Methanocaldococcus Jannaschii Escherichia Coli E. ColiFederico VillanuevaPas encore d'évaluation
- Molecular Pharmacology Exam Questions With AnswersDocument12 pagesMolecular Pharmacology Exam Questions With AnswersAdam McLaren100% (1)
- CH12 159-168 PDFDocument10 pagesCH12 159-168 PDFidjacobsPas encore d'évaluation
- EPILEPSY Part 2Document30 pagesEPILEPSY Part 2abdulPas encore d'évaluation
- Questions To Answer:: Effects On The BrainDocument7 pagesQuestions To Answer:: Effects On The BrainJade Phoebe AjeroPas encore d'évaluation
- Chapter Sixteen Cell SignalingDocument96 pagesChapter Sixteen Cell SignalingRu LiliPas encore d'évaluation
- J. Biol. Chem.-2012-Sigel-40224-31 PDFDocument9 pagesJ. Biol. Chem.-2012-Sigel-40224-31 PDFIchim VladPas encore d'évaluation
- CalcioDocument6 pagesCalcioArtur Franck Tenorio AronésPas encore d'évaluation
- Andreychenko Et Al. - 2012 - Efficient Spectral Editing at 7 T GABA Detection With MEGA-sLASERDocument8 pagesAndreychenko Et Al. - 2012 - Efficient Spectral Editing at 7 T GABA Detection With MEGA-sLASERjan5437Pas encore d'évaluation
- Receptors Ion ChannelsDocument42 pagesReceptors Ion ChannelsMd. Ahsan-Ul BariPas encore d'évaluation
- Neurochem FinalDocument20 pagesNeurochem Finalelifulku.a1Pas encore d'évaluation
- GABAB Receptors and DiseaseDocument5 pagesGABAB Receptors and Diseaselucia desantisPas encore d'évaluation
- Current Concepts in The Biochemical Mechanisms of Glaucomatous NeurodegenerationDocument0 pageCurrent Concepts in The Biochemical Mechanisms of Glaucomatous NeurodegenerationlovehopePas encore d'évaluation
- Lab 5 and 6Document4 pagesLab 5 and 6Erika Mae EspejoPas encore d'évaluation
- Principles of Drug Action RevisedDocument44 pagesPrinciples of Drug Action RevisedRachma AthayaPas encore d'évaluation
- BIOCHEMISTRY NotesDocument127 pagesBIOCHEMISTRY NotesMelanie Brittain100% (1)
- Lecture Notes Urea Cycle Biochemistry 2, 2016 FV: Non-Essential Amino Acids Nitrogen MetabolismDocument34 pagesLecture Notes Urea Cycle Biochemistry 2, 2016 FV: Non-Essential Amino Acids Nitrogen MetabolismChaitali Dongaonkar ChawarePas encore d'évaluation
- Presentation On CGMP AccumulationDocument1 pagePresentation On CGMP Accumulationhi hiPas encore d'évaluation
- B22 Neural Mechanisms of Learning & MemoryDocument21 pagesB22 Neural Mechanisms of Learning & MemoryNilanjana Chatterjee ChakrabortyPas encore d'évaluation
- Aula 7 Comunicao Celular-farm-22.2.XxxDocument51 pagesAula 7 Comunicao Celular-farm-22.2.XxxPersonal Trainer Rafael Cruz do NascimentoPas encore d'évaluation
- EpilepsyDocument6 pagesEpilepsylucia desantisPas encore d'évaluation
- KrebcycleDocument5 pagesKrebcycleYana PajoPas encore d'évaluation
- NHE3 ZIjian XieDocument9 pagesNHE3 ZIjian Xienilberto2Pas encore d'évaluation
- Gaba Is The Major Inhibitory Neurotransmitter in The Nervous SystemDocument10 pagesGaba Is The Major Inhibitory Neurotransmitter in The Nervous SystemMount EmeraldPas encore d'évaluation
- 2022 Mechanisms of Inhibition and Activation of Extrasynaptic Ab GABAa Receptors - CompressedDocument23 pages2022 Mechanisms of Inhibition and Activation of Extrasynaptic Ab GABAa Receptors - CompressedVitoria LimaPas encore d'évaluation
- Receptor SuperfamiliesDocument27 pagesReceptor SuperfamiliesNAUSHEEN SULTANAPas encore d'évaluation
- Informe Práctica de Laboratorio 6 - Respuesta en Frecuencia y ResonanciaDocument11 pagesInforme Práctica de Laboratorio 6 - Respuesta en Frecuencia y Resonanciaeric2717Pas encore d'évaluation
- Electrophysiology and Electro Physio Pharmacology of Cardiac Cells - Docclass 2 Electrophysiology and Electro Physio Pharmacology of Cardiac CellsDocument71 pagesElectrophysiology and Electro Physio Pharmacology of Cardiac Cells - Docclass 2 Electrophysiology and Electro Physio Pharmacology of Cardiac CellsdcicarePas encore d'évaluation
- s41467 019 13766 2 PDFDocument11 pagess41467 019 13766 2 PDFCarolina RicárdezPas encore d'évaluation
- NPB 100 Midterm 1 Study Guide-1Document4 pagesNPB 100 Midterm 1 Study Guide-1April MartínezPas encore d'évaluation
- Sodium Channel Structure, Function, Gating and Involvement in DiseaseDocument38 pagesSodium Channel Structure, Function, Gating and Involvement in DiseaseYelsa NoritaPas encore d'évaluation
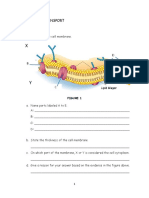
- Cell Transport Review WorksheetsDocument8 pagesCell Transport Review WorksheetsD50% (2)
- Chapter 4Document44 pagesChapter 4LynettraPas encore d'évaluation
- 2 5 Transport MechanismDocument25 pages2 5 Transport Mechanismıamnıkolaı 4100% (1)
- Neuronal Conduction1Document61 pagesNeuronal Conduction1ISHITA GOEL 19224115Pas encore d'évaluation
- Defining Solution Conformations of Small Linear PeptidesDocument20 pagesDefining Solution Conformations of Small Linear PeptidesSevim KösePas encore d'évaluation
- 2D NMR Spectroscopy - 1-D and 2-D NMR, Noesy and Cosy, Hetcor, INADEQUATE TechniquesDocument21 pages2D NMR Spectroscopy - 1-D and 2-D NMR, Noesy and Cosy, Hetcor, INADEQUATE TechniquesАрсений БекишевPas encore d'évaluation
- Nonspecific and Specific DefensesDocument3 pagesNonspecific and Specific DefensesLorenz Jude CańetePas encore d'évaluation
- QUITALIG Biochemistry Lecture Module 5 ActivityDocument5 pagesQUITALIG Biochemistry Lecture Module 5 ActivityAloysius QuitaligPas encore d'évaluation
- Crystal Structure - Part 1Document14 pagesCrystal Structure - Part 1Martin VailPas encore d'évaluation
- Liposome Drug Delivery SystemDocument28 pagesLiposome Drug Delivery SystemGANESH KUMAR JELLA100% (1)
- Energy: Chapter 5 - The Working CellDocument26 pagesEnergy: Chapter 5 - The Working CellJohn Paul Coronel SinuhinPas encore d'évaluation
- Primitive Lattice Vectors (Primitive Translation Vectors)Document34 pagesPrimitive Lattice Vectors (Primitive Translation Vectors)VarshaPas encore d'évaluation
- Term-Project-Ii-Bch3125 ADocument8 pagesTerm-Project-Ii-Bch3125 APure PurePas encore d'évaluation
- Membrane PotentialsDocument40 pagesMembrane PotentialsdehaPas encore d'évaluation
- Unit IV CrystallographyDocument17 pagesUnit IV Crystallographyswapniljawadwar1Pas encore d'évaluation
- Reciprocal Lattice & Brillouin ZoneDocument20 pagesReciprocal Lattice & Brillouin ZoneAkshay KumarPas encore d'évaluation
- Overview of The Immune System: With Prof. Peter J. DelvesDocument27 pagesOverview of The Immune System: With Prof. Peter J. Delveslow ah keongPas encore d'évaluation
- Crystal Structure and Crystal GeometryDocument103 pagesCrystal Structure and Crystal GeometryKhalid Al ShaaibiPas encore d'évaluation
- Su 1 Le 1 - 1Document18 pagesSu 1 Le 1 - 1Francisca ManyisaPas encore d'évaluation
- Solutions MidtermDocument3 pagesSolutions MidtermAli NajmaldinPas encore d'évaluation
- Cellular Transport PowerpointDocument50 pagesCellular Transport PowerpointPortia EgkenPas encore d'évaluation
- Chapter 1 Elementary Materials Science PDFDocument33 pagesChapter 1 Elementary Materials Science PDFSohan HasanPas encore d'évaluation
- The Solid State Notes PDFDocument37 pagesThe Solid State Notes PDFSandy 05Pas encore d'évaluation
- TOPIC 3 - Cell TransportDocument7 pagesTOPIC 3 - Cell TransportSiti Nurqistina Balqis Syahrul EffendyPas encore d'évaluation
- Cell Transport PowerpointDocument32 pagesCell Transport PowerpointtanyaPas encore d'évaluation
- Calcium As A Secondary Messenger in Cell SignallingDocument5 pagesCalcium As A Secondary Messenger in Cell Signallingjeanoriordan02Pas encore d'évaluation
- Reciprocal Lattice & Ewald Sphere ConstructionDocument45 pagesReciprocal Lattice & Ewald Sphere ConstructionSplendid SciencePas encore d'évaluation
- 1.05 SphingolipidsDocument5 pages1.05 Sphingolipidslearningp18Pas encore d'évaluation
- Intercellular Junctions: Alan S. L. Yu, Fiona Hanner and Ja Nos Peti-PeterdiDocument1 pageIntercellular Junctions: Alan S. L. Yu, Fiona Hanner and Ja Nos Peti-PeterdiaaaatniPas encore d'évaluation
- Cardiac Action PotentialDocument5 pagesCardiac Action PotentialjayPas encore d'évaluation
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsD'EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsPas encore d'évaluation
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDD'EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDÉvaluation : 5 sur 5 étoiles5/5 (3)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionD'EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionÉvaluation : 4 sur 5 étoiles4/5 (404)
- The Age of Magical Overthinking: Notes on Modern IrrationalityD'EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityÉvaluation : 4 sur 5 étoiles4/5 (32)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeD'EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeÉvaluation : 2 sur 5 étoiles2/5 (1)
- The Comfort of Crows: A Backyard YearD'EverandThe Comfort of Crows: A Backyard YearÉvaluation : 4.5 sur 5 étoiles4.5/5 (23)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)D'EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Évaluation : 3 sur 5 étoiles3/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedD'EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedÉvaluation : 4.5 sur 5 étoiles4.5/5 (82)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsD'EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsÉvaluation : 4 sur 5 étoiles4/5 (4)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisD'EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisÉvaluation : 4.5 sur 5 étoiles4.5/5 (42)
- The Obesity Code: Unlocking the Secrets of Weight LossD'EverandThe Obesity Code: Unlocking the Secrets of Weight LossÉvaluation : 4 sur 5 étoiles4/5 (6)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsD'EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessD'EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessÉvaluation : 4.5 sur 5 étoiles4.5/5 (328)
- To Explain the World: The Discovery of Modern ScienceD'EverandTo Explain the World: The Discovery of Modern ScienceÉvaluation : 3.5 sur 5 étoiles3.5/5 (51)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaD'EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesD'EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesÉvaluation : 4.5 sur 5 étoiles4.5/5 (1412)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryD'EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryÉvaluation : 4 sur 5 étoiles4/5 (46)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeD'EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeÉvaluation : 4.5 sur 5 étoiles4.5/5 (254)