Vous aimerez peut-être aussi
- House 2003Document16 pagesHouse 2003sandrino1091Pas encore d'évaluation
- Allometry of Aboveground Biomasses in Mangrove Species in Itamaraca, Pernambuco, BrazilDocument8 pagesAllometry of Aboveground Biomasses in Mangrove Species in Itamaraca, Pernambuco, BrazilDaniel PaivaPas encore d'évaluation
- Importance of Structure and Its Measurement in Quantifying Function of Forest EcosystemsDocument16 pagesImportance of Structure and Its Measurement in Quantifying Function of Forest EcosystemsRodrigo vazquez moralesPas encore d'évaluation
- 8104 PDFDocument12 pages8104 PDFRahul JerryPas encore d'évaluation
- NZJEcol38 1 ThrippletonIP SuppDocument3 pagesNZJEcol38 1 ThrippletonIP SuppioaPas encore d'évaluation
- LAI Calc DiameteratbreastDocument12 pagesLAI Calc Diameteratbreastjohnnypie7Pas encore d'évaluation
- P3nting 2Document7 pagesP3nting 2Putranto AgungPas encore d'évaluation
- Species Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringDocument13 pagesSpecies Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringLight WhitePas encore d'évaluation
- Ecological Monographs - 2004 - Ackerly - FUNCTIONAL STRATEGIES OF CHAPARRAL SHRUBS IN RELATION TO SEASONAL WATER DEFICITDocument20 pagesEcological Monographs - 2004 - Ackerly - FUNCTIONAL STRATEGIES OF CHAPARRAL SHRUBS IN RELATION TO SEASONAL WATER DEFICITAna SampaioPas encore d'évaluation
- Carlucci2012 - Tree Traits (Análises)Document11 pagesCarlucci2012 - Tree Traits (Análises)BeatrizPas encore d'évaluation
- Aparecido2019 - SapwoodDocument15 pagesAparecido2019 - SapwoodcristinePas encore d'évaluation
- Foliage Density of Branches and Distribution of Plant Dwelling SpidersDocument5 pagesFoliage Density of Branches and Distribution of Plant Dwelling SpidersCélio Moura NetoPas encore d'évaluation
- CSR Analysis of Plant Functional Types in Highly Diverse Tropical Grasslands of Harsh EnvironmentsDocument10 pagesCSR Analysis of Plant Functional Types in Highly Diverse Tropical Grasslands of Harsh EnvironmentsGuilherme FerreiraPas encore d'évaluation
- Relevance of Wood Anatomy and Size Amazonian Trees in The Determination and Allometry of Sapwood AreaDocument16 pagesRelevance of Wood Anatomy and Size Amazonian Trees in The Determination and Allometry of Sapwood AreaAlejandro Guerrero LaverdePas encore d'évaluation
- Goetz 2014Document19 pagesGoetz 2014Daniele SoardiPas encore d'évaluation
- Vegetation Map - BINP - Babaasa Et Al. 2024Document13 pagesVegetation Map - BINP - Babaasa Et Al. 2024BabaasaPas encore d'évaluation
- Gough C Et Al 2019 EcologyDocument6 pagesGough C Et Al 2019 EcologyGiorgio VacchianoPas encore d'évaluation
- Modeling Forest Canopy Structure and Density by Combining Point Quadrat Sampling and Survival AnalysisDocument12 pagesModeling Forest Canopy Structure and Density by Combining Point Quadrat Sampling and Survival AnalysisAldo QuesadaPas encore d'évaluation
- Use of Boundary-Line Growth Patterns As A Basis For Dendroecological Release CriteriaDocument17 pagesUse of Boundary-Line Growth Patterns As A Basis For Dendroecological Release CriteriaCatalin RoibuPas encore d'évaluation
- Sucesion Del BosueDocument11 pagesSucesion Del BosueMile PenagosPas encore d'évaluation
- Lu Et Al 2003 Sucesional Stages PDFDocument12 pagesLu Et Al 2003 Sucesional Stages PDFVanessa AraujoPas encore d'évaluation
- Assessing Impact of Projected Future ClimateDocument15 pagesAssessing Impact of Projected Future ClimateGuillermo EsparzaPas encore d'évaluation
- COOMES Et Al-2007-Journal of EcologyDocument14 pagesCOOMES Et Al-2007-Journal of EcologyRajeev DhimanPas encore d'évaluation
- López 2010 Patrones de Diversidad Alfa en Tres Fragmentos de Bosques MontanosDocument16 pagesLópez 2010 Patrones de Diversidad Alfa en Tres Fragmentos de Bosques MontanosDaniela Vasquez VergaraPas encore d'évaluation
- Morpologi Dan Pollen DispersalDocument11 pagesMorpologi Dan Pollen DispersalsecrettasteuhhPas encore d'évaluation
- FullDocument50 pagesFullabidah fitraPas encore d'évaluation
- 2018maxdensity EcoEvolDocument12 pages2018maxdensity EcoEvolChristian Salas-EljatibPas encore d'évaluation
- Mattia Et La., 2005Document10 pagesMattia Et La., 2005Pawan SharmaPas encore d'évaluation
- 2018 Rodrigues Et Al Plant Ecol DivDocument12 pages2018 Rodrigues Et Al Plant Ecol DivEmerson PawoskiPas encore d'évaluation
- Mensah Et Al. 2016b - Biomass Allocation Patterns - Annals of Forest ResearchDocument12 pagesMensah Et Al. 2016b - Biomass Allocation Patterns - Annals of Forest ResearchMensah sylvanusPas encore d'évaluation
- Variacion RADIAL de DensidadDocument7 pagesVariacion RADIAL de DensidadErnesto CoparaPas encore d'évaluation
- Bosque Lluvioso - 02Document10 pagesBosque Lluvioso - 02yuranyPas encore d'évaluation
- Assessment of Biomass Content and Oxygen Production From Tree of Subtropical AreaDocument5 pagesAssessment of Biomass Content and Oxygen Production From Tree of Subtropical AreaResearch ParkPas encore d'évaluation
- Jadan Et Al 2022 Forest EcosystemsDocument9 pagesJadan Et Al 2022 Forest EcosystemsOswaldo JadanPas encore d'évaluation
- O62F3754Document216 pagesO62F3754jegarciapPas encore d'évaluation
- Belote Et Al. 2009Document8 pagesBelote Et Al. 2009Carla VitorinoPas encore d'évaluation
- Plant Functional Groups Within A Tropical Forest Exhibit Different Wood Functional AnatomyDocument10 pagesPlant Functional Groups Within A Tropical Forest Exhibit Different Wood Functional AnatomyAriadne Cristina De AntonioPas encore d'évaluation
- Noreika & Kotze - 2012Document15 pagesNoreika & Kotze - 2012Dalius DapkusPas encore d'évaluation
- Rodríguez-Alarcón2018 Article EffectsOfFragmentationOnFunctiDocument14 pagesRodríguez-Alarcón2018 Article EffectsOfFragmentationOnFunctitmgutierrez14Pas encore d'évaluation
- Muñoz Mazón Et Al., 2020Document9 pagesMuñoz Mazón Et Al., 2020azeta zetaPas encore d'évaluation
- Discussion Adult SeedelingDocument13 pagesDiscussion Adult SeedelingAlejandra LlerenaPas encore d'évaluation
- Volume Models ForestryDocument12 pagesVolume Models ForestryStalin FernandezPas encore d'évaluation
- Classification of Successional Forest Stages in The Brazilian Amazon BasinDocument12 pagesClassification of Successional Forest Stages in The Brazilian Amazon BasinTytyt TyyPas encore d'évaluation
- Urbina-Cardona Et Al 2006 BiolconsDocument15 pagesUrbina-Cardona Et Al 2006 BiolconsDaniel VásquezPas encore d'évaluation
- Tuomisto Et Al 2003 - ScienceDocument5 pagesTuomisto Et Al 2003 - ScienceKátia FernandaPas encore d'évaluation
- Tree Physiol 1991 Milne 383 99Document18 pagesTree Physiol 1991 Milne 383 99vfanghelinaPas encore d'évaluation
- Coley 1988Document6 pagesColey 1988Gabriela MouraPas encore d'évaluation
- Tree Ring Analysis of Mexican Weeping Pine (Pinus Patula Schiede Ex Schltdl. and Cham.) From Nagaland, Northeast IndiaDocument11 pagesTree Ring Analysis of Mexican Weeping Pine (Pinus Patula Schiede Ex Schltdl. and Cham.) From Nagaland, Northeast Indiasmiling01011980Pas encore d'évaluation
- Zekeng Et Al 2023Document13 pagesZekeng Et Al 2023Citoyen x14Pas encore d'évaluation
- Diversity and Big Trees Patterns in The Brazilian Amazon.Document10 pagesDiversity and Big Trees Patterns in The Brazilian Amazon.Cindy OlivierPas encore d'évaluation
- Multiple Edge Effects and Their Implications in Fragmented LandscapesDocument11 pagesMultiple Edge Effects and Their Implications in Fragmented LandscapesIngForestalPas encore d'évaluation
- Aldrich 1998Document12 pagesAldrich 1998Rany sanchium tawamPas encore d'évaluation
- Mensah Et Al. 2018b Differential Responses of TaxonomicDocument13 pagesMensah Et Al. 2018b Differential Responses of TaxonomicMensah sylvanusPas encore d'évaluation
- Edge-Interior Differences in Vegetation Structure and Composition in A Chaco Semi-Arid Forest, ArgentinaDocument9 pagesEdge-Interior Differences in Vegetation Structure and Composition in A Chaco Semi-Arid Forest, ArgentinaAnonymous ZON7qMPas encore d'évaluation
- Duran Et Al 2015 The Relative Importance of Climate, Stand Variables and Liana Abundance For Carbon Storage in Tropical Forests Climate, Stand Variables and Lianas Control Forest CarbonDocument12 pagesDuran Et Al 2015 The Relative Importance of Climate, Stand Variables and Liana Abundance For Carbon Storage in Tropical Forests Climate, Stand Variables and Lianas Control Forest Carbonfitri.ainiPas encore d'évaluation
- Plant Strategie1Document6 pagesPlant Strategie1Shahzadali Muhammad ibrahimPas encore d'évaluation
- Chen 1995Document14 pagesChen 1995Ryan MarquesPas encore d'évaluation
- 70845-Texto Del Artículo-423490-3-10-20190708Document12 pages70845-Texto Del Artículo-423490-3-10-20190708WILLIAM ENRIQUE FIGUEROA MARTINEZPas encore d'évaluation
- Growth-Ring Boundary Anatomy and Dendrochronological Potential in A Moist Tropical Forest in Northeastern BangladeshDocument19 pagesGrowth-Ring Boundary Anatomy and Dendrochronological Potential in A Moist Tropical Forest in Northeastern BangladeshAnthony GuerraPas encore d'évaluation
- Applications of Physiological Ecology to Forest ManagementD'EverandApplications of Physiological Ecology to Forest ManagementPas encore d'évaluation
- Dendrochronologia: Anthony M. Fowler, Martin C. BridgeDocument5 pagesDendrochronologia: Anthony M. Fowler, Martin C. BridgeCatalin RoibuPas encore d'évaluation
- Dendrochronologia: Katarina Cufar, Matja Z Bizjak, Manja Kitek Kuzman, Maks Merela, Michael Grabner, Robert BrusDocument7 pagesDendrochronologia: Katarina Cufar, Matja Z Bizjak, Manja Kitek Kuzman, Maks Merela, Michael Grabner, Robert BrusCatalin RoibuPas encore d'évaluation
- Manuscript 40329 1 10 20161214Document9 pagesManuscript 40329 1 10 20161214Catalin RoibuPas encore d'évaluation
- Type Metric Adjusted R: Mean Age3 Age5 AGE10Document1 pageType Metric Adjusted R: Mean Age3 Age5 AGE10Catalin RoibuPas encore d'évaluation
- Table 2.: Table S3. Raw Values of The Selected Metrics Used in The Analyses. Variable Codes Explained inDocument1 pageTable 2.: Table S3. Raw Values of The Selected Metrics Used in The Analyses. Variable Codes Explained inCatalin RoibuPas encore d'évaluation
- Package Treeclim': October 8, 2020Document23 pagesPackage Treeclim': October 8, 2020Catalin RoibuPas encore d'évaluation
- Forestry: A Review of European Ash (FraxinusDocument16 pagesForestry: A Review of European Ash (FraxinusCatalin RoibuPas encore d'évaluation
- Figure S2. Comparison Between The Loadings On The First Principal Component (PC1) of All The 39 ChronoDocument1 pageFigure S2. Comparison Between The Loadings On The First Principal Component (PC1) of All The 39 ChronoCatalin RoibuPas encore d'évaluation
- Jpe12793 Sup 0008 TableS2Document1 pageJpe12793 Sup 0008 TableS2Catalin RoibuPas encore d'évaluation
- Jpe12793 Sup 0010 TableS4Document1 pageJpe12793 Sup 0010 TableS4Catalin RoibuPas encore d'évaluation
- Figure S1. Growth Trajectories Within Beech Forests of The Apennines (Above) and The Eastern Alps (Below)Document1 pageFigure S1. Growth Trajectories Within Beech Forests of The Apennines (Above) and The Eastern Alps (Below)Catalin RoibuPas encore d'évaluation
- Jpe12793 Sup 0010 TableS4Document1 pageJpe12793 Sup 0010 TableS4Catalin RoibuPas encore d'évaluation
- A S2. Suppression CalculationDocument3 pagesA S2. Suppression CalculationCatalin RoibuPas encore d'évaluation
- Jpe12793 Sup 0001 AppendixS1Document7 pagesJpe12793 Sup 0001 AppendixS1Catalin RoibuPas encore d'évaluation
- Lifetime Growth Patterns and Ages of Bolivian Rainforest Trees Obtained by Tree Ring AnalysisDocument18 pagesLifetime Growth Patterns and Ages of Bolivian Rainforest Trees Obtained by Tree Ring AnalysisCatalin RoibuPas encore d'évaluation
- Use of Boundary-Line Growth Patterns As A Basis For Dendroecological Release CriteriaDocument17 pagesUse of Boundary-Line Growth Patterns As A Basis For Dendroecological Release CriteriaCatalin RoibuPas encore d'évaluation
- Art:10.1007/s00382 008 0395 5 PDFDocument15 pagesArt:10.1007/s00382 008 0395 5 PDFCatalin RoibuPas encore d'évaluation
- Statistical Ecology. A Primer On Methods and ComputingDocument358 pagesStatistical Ecology. A Primer On Methods and ComputingCatalin RoibuPas encore d'évaluation
- Novedades Jaltest CV en 887Document14 pagesNovedades Jaltest CV en 887Bruce LyndePas encore d'évaluation
- Educ 3 Prelim Act.1 AlidonDocument2 pagesEduc 3 Prelim Act.1 AlidonJonash AlidonPas encore d'évaluation
- Stress Corrosion Cracking Behavior of X80 PipelineDocument13 pagesStress Corrosion Cracking Behavior of X80 Pipelineaashima sharmaPas encore d'évaluation
- The Neurology of Ageing: What Is NormalDocument11 pagesThe Neurology of Ageing: What Is NormalEsther EunikePas encore d'évaluation
- The Permission Wheel Hawkes L PDFDocument8 pagesThe Permission Wheel Hawkes L PDFNandita Asthana SankerPas encore d'évaluation
- Mendoza - Kyle Andre - BSEE-1A (STS ACTIVITY 5)Document1 pageMendoza - Kyle Andre - BSEE-1A (STS ACTIVITY 5)Kyle Andre MendozaPas encore d'évaluation
- KV3000 Programming TutorialDocument42 pagesKV3000 Programming TutorialanthonyPas encore d'évaluation
- Land Building and MachineryDocument26 pagesLand Building and MachineryNathalie Getino100% (1)
- Marriot CaseDocument15 pagesMarriot CaseArsh00100% (7)
- KB000120-MRK456-01-HR SamplingDocument15 pagesKB000120-MRK456-01-HR SamplingMiguel Zuniga MarconiPas encore d'évaluation
- Durst Caldera: Application GuideDocument70 pagesDurst Caldera: Application GuideClaudio BasconiPas encore d'évaluation
- 19-Microendoscopic Lumbar DiscectomyDocument8 pages19-Microendoscopic Lumbar DiscectomyNewton IssacPas encore d'évaluation
- Advanced Financial Accounting and Reporting Accounting For PartnershipDocument6 pagesAdvanced Financial Accounting and Reporting Accounting For PartnershipMaria BeatricePas encore d'évaluation
- The Leaders of The NationDocument3 pagesThe Leaders of The NationMark Dave RodriguezPas encore d'évaluation
- Wastewater Treatment: Sudha Goel, Ph.D. Department of Civil Engineering, IIT KharagpurDocument33 pagesWastewater Treatment: Sudha Goel, Ph.D. Department of Civil Engineering, IIT KharagpurSubhajit BagPas encore d'évaluation
- Book TurmericDocument14 pagesBook Turmericarvind3041990100% (2)
- Psychology Research Literature Review ExampleDocument5 pagesPsychology Research Literature Review Exampleafdtsebxc100% (1)
- Answer Key For 1st QaurterDocument5 pagesAnswer Key For 1st QaurterSteffi89% (9)
- Human Development IndexDocument17 pagesHuman Development IndexriyaPas encore d'évaluation
- TTPQDocument2 pagesTTPQchrystal85Pas encore d'évaluation
- Science Technology and SocietyDocument46 pagesScience Technology and SocietyCharles Elquime GalaponPas encore d'évaluation
- Literatures of The World: Readings For Week 4 in LIT 121Document11 pagesLiteratures of The World: Readings For Week 4 in LIT 121April AcompaniadoPas encore d'évaluation
- Health and Hatha Yoga by Swami Sivananda CompressDocument356 pagesHealth and Hatha Yoga by Swami Sivananda CompressLama Fera with Yachna JainPas encore d'évaluation
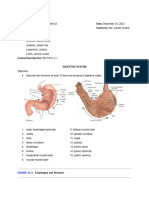
- Digestive System LabsheetDocument4 pagesDigestive System LabsheetKATHLEEN MAE HERMOPas encore d'évaluation
- Iluminadores y DipolosDocument9 pagesIluminadores y DipolosRamonPas encore d'évaluation
- DE1734859 Central Maharashtra Feb'18Document39 pagesDE1734859 Central Maharashtra Feb'18Adesh NaharPas encore d'évaluation
- MagmatismDocument12 pagesMagmatismVea Patricia Angelo100% (1)
- UXBenchmarking 101Document42 pagesUXBenchmarking 101Rodrigo BucketbranchPas encore d'évaluation
- ANSI-ISA-S5.4-1991 - Instrument Loop DiagramsDocument22 pagesANSI-ISA-S5.4-1991 - Instrument Loop DiagramsCarlos Poveda100% (2)
- Supply Chain Management: A Framework of Understanding D. Du Toit & P.J. VlokDocument14 pagesSupply Chain Management: A Framework of Understanding D. Du Toit & P.J. VlokchandanaPas encore d'évaluation