Vous aimerez peut-être aussi
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Carpal Tunnel Syndrome - Kiiko Matsumoto Japanese StyleDocument3 pagesCarpal Tunnel Syndrome - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Allergies - Kiiko Matsumoto Japanese StyleDocument4 pagesAllergies - Kiiko Matsumoto Japanese StyleAWEDIOHEAD100% (1)
- Cough - Kiiko Matsumoto Japanese StyleDocument3 pagesCough - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Depression - Kiiko Matsumoto Japanese StyleDocument3 pagesDepression - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Constipation - Kiiko Matsumoto Japanese StyleDocument2 pagesConstipation - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Asthma - Kiiko Matsumoto Japanese StyleDocument4 pagesAsthma - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Abnormal Menstrual Bleeding - Kiiko Matsumoto Japanese StyleDocument3 pagesAbnormal Menstrual Bleeding - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Abdominal Distention and Bloating - Kiiko Matsumoto Japanese StyleDocument4 pagesAbdominal Distention and Bloating - Kiiko Matsumoto Japanese StyleAWEDIOHEAD100% (2)
- Anxiety - Kiiko Matsumoto Japanese StyleDocument5 pagesAnxiety - Kiiko Matsumoto Japanese StyleAWEDIOHEADPas encore d'évaluation
- Router Set-Up Guide - Technicolor TG588 in Bridge Modem ModeDocument6 pagesRouter Set-Up Guide - Technicolor TG588 in Bridge Modem ModeAWEDIOHEADPas encore d'évaluation
- Notes On IGCSE English PoemsDocument32 pagesNotes On IGCSE English Poemsbluehorse99Pas encore d'évaluation
- Hormonal ImbalanceDocument1 pageHormonal ImbalanceAWEDIOHEAD100% (9)
- Culture Techniques of Moina The Ideal Daphnia For Feeding Freshwater Fish Fry1 PDFDocument7 pagesCulture Techniques of Moina The Ideal Daphnia For Feeding Freshwater Fish Fry1 PDFPerpetua KamikadzePas encore d'évaluation
- Metal Shed Assembly InstructionsDocument29 pagesMetal Shed Assembly InstructionsAWEDIOHEADPas encore d'évaluation
- Lu Xu Yang Excess HeatDocument2 pagesLu Xu Yang Excess HeatAWEDIOHEADPas encore d'évaluation
- AdrenalsDocument1 pageAdrenalsAWEDIOHEADPas encore d'évaluation
- Acupunture Thought English PDFDocument56 pagesAcupunture Thought English PDFAWEDIOHEAD100% (8)
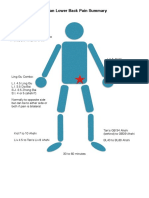
- DR Tan Low Back Pain SummaryDocument1 pageDR Tan Low Back Pain SummaryAWEDIOHEAD92% (13)
- A Step by Step Guide To Dirting Your TankDocument4 pagesA Step by Step Guide To Dirting Your TankAWEDIOHEADPas encore d'évaluation
- Questions To Senior AcupuncturistsDocument15 pagesQuestions To Senior AcupuncturistsAWEDIOHEAD100% (2)
- Master Tung's Four Horses Combination - Master Tung's Magic PointsDocument12 pagesMaster Tung's Four Horses Combination - Master Tung's Magic PointsAWEDIOHEAD93% (14)
- Ikeda Points NotesDocument1 pageIkeda Points NotesAWEDIOHEADPas encore d'évaluation
- Birch - Contraindications in PregnancyDocument10 pagesBirch - Contraindications in PregnancyAWEDIOHEAD100% (2)
- Birch - German Acupuncture ResearchDocument6 pagesBirch - German Acupuncture ResearchAWEDIOHEADPas encore d'évaluation
- HaraShiatsu Self HealingDocument3 pagesHaraShiatsu Self HealingAWEDIOHEADPas encore d'évaluation
- Ondan Guards Designed To Be Used in Pairs. Print On Thick Card With A "Borderless" Setting IdeallyDocument1 pageOndan Guards Designed To Be Used in Pairs. Print On Thick Card With A "Borderless" Setting IdeallyAWEDIOHEADPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Defining and Non-Defining ClausesDocument14 pagesDefining and Non-Defining ClausesАнастасия РезановаPas encore d'évaluation
- Molecular Plant Taxonomy. Methods in Molecular Biology (2021)Document407 pagesMolecular Plant Taxonomy. Methods in Molecular Biology (2021)Giovanni Celso Paton MamaniPas encore d'évaluation
- Nta Neet Abhyas MOCK TEST - 1Document40 pagesNta Neet Abhyas MOCK TEST - 1bapiPas encore d'évaluation
- University of Cambridge International Examinations International General Certificate of Secondary EducationDocument20 pagesUniversity of Cambridge International Examinations International General Certificate of Secondary EducationShivraj NaikPas encore d'évaluation
- Homo Sapiens Chromosome 8, GRCh38.p13 Primary Assembly - Nucleotide - NCBIDocument1 pageHomo Sapiens Chromosome 8, GRCh38.p13 Primary Assembly - Nucleotide - NCBIkameliyaPas encore d'évaluation
- Pepeperiksaan Pra-STPM Penggal 3 2017 (Soalan)Document7 pagesPepeperiksaan Pra-STPM Penggal 3 2017 (Soalan)Viola Voon Li WeiPas encore d'évaluation
- Down SyndromeDocument17 pagesDown SyndromeSarah Shine TorresPas encore d'évaluation
- Neurons and Nerve Impulses: Nandika Arora and Risa Gaikwad (11 G2)Document17 pagesNeurons and Nerve Impulses: Nandika Arora and Risa Gaikwad (11 G2)RisaPas encore d'évaluation
- Danger Avoidance Can Be Genetically Encoded For Four Generations, Say Biologists - Scientists Found That Learned Behaviors in CDocument3 pagesDanger Avoidance Can Be Genetically Encoded For Four Generations, Say Biologists - Scientists Found That Learned Behaviors in Cdamdidam1Pas encore d'évaluation
- Lec-Bio103L, Expt 6 Summer 2023-1Document9 pagesLec-Bio103L, Expt 6 Summer 2023-1Maysha RodoshiPas encore d'évaluation
- Polymerase Chain Reaction (PCR) (Article) - Khan AcademyDocument10 pagesPolymerase Chain Reaction (PCR) (Article) - Khan AcademySelva KumarPas encore d'évaluation
- Cell and Molecular Biology - What We KnowDocument11 pagesCell and Molecular Biology - What We Knowhar harPas encore d'évaluation
- Microbiology Control: Techniques For Controlling Pathogenic MicroorganismDocument22 pagesMicrobiology Control: Techniques For Controlling Pathogenic MicroorganismEunice MañalacPas encore d'évaluation
- Fand 2019Document13 pagesFand 2019Vishnuvardhan RavichandranPas encore d'évaluation
- Polyplus Transfection® Life Science Research and Non-Viral In-Vivo Transfection ReagentsDocument6 pagesPolyplus Transfection® Life Science Research and Non-Viral In-Vivo Transfection ReagentsDiego GarridoPas encore d'évaluation
- All India Test Series (Neet) : RMCT-01ADocument14 pagesAll India Test Series (Neet) : RMCT-01AManmeet SinghPas encore d'évaluation
- Hematology I - Hemoglobin: Physiologic FormsDocument4 pagesHematology I - Hemoglobin: Physiologic FormsAldren BeliberPas encore d'évaluation
- Test Bank For Exploring Psychology in Modules, 12e David Myers, Nathan DeWall Test BankDocument34 pagesTest Bank For Exploring Psychology in Modules, 12e David Myers, Nathan DeWall Test BankNail BaskoPas encore d'évaluation
- Research Proposal-Kelsi GelleDocument5 pagesResearch Proposal-Kelsi Gelleapi-619041501Pas encore d'évaluation
- 1 s2.0 S1568988322000166 MainDocument12 pages1 s2.0 S1568988322000166 MainAlexandru IorgescuPas encore d'évaluation
- GEARY, Patrick. Genetic History and Migration in Western Eurasia (500-1000), em DIDocument16 pagesGEARY, Patrick. Genetic History and Migration in Western Eurasia (500-1000), em DILavinia AnjosPas encore d'évaluation
- Test Report: MR - Raghavan Venkatraman (39/M)Document2 pagesTest Report: MR - Raghavan Venkatraman (39/M)Raghavan VenkatramanPas encore d'évaluation
- The Studying The Biological Characteristics of The Stingless Bee Tetrigonilla Collina Smith 1857Document10 pagesThe Studying The Biological Characteristics of The Stingless Bee Tetrigonilla Collina Smith 1857dphvuPas encore d'évaluation
- WBI01 01 Rms 20190124Document28 pagesWBI01 01 Rms 20190124Adrian Safa khouryPas encore d'évaluation
- Navneet QB - Science Grade 10Document36 pagesNavneet QB - Science Grade 10Himanshu NagarePas encore d'évaluation
- ) "PDF PDFDocument18 pages) "PDF PDFRyan LiPas encore d'évaluation
- 08 Worksheet 2Document9 pages08 Worksheet 2James TevesPas encore d'évaluation
- Activity 1.5cell DivisionDocument2 pagesActivity 1.5cell Divisionfroilan obligadoPas encore d'évaluation
- Lecture 1 Nature and Scope of Developmental Biology (Part I)Document34 pagesLecture 1 Nature and Scope of Developmental Biology (Part I)Zari Sofia LevistePas encore d'évaluation
- WHO - Active Applications - Covid Testing - EULDocument6 pagesWHO - Active Applications - Covid Testing - EULbharat1102Pas encore d'évaluation