Vous aimerez peut-être aussi
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Epic TutDocument15 pagesEpic TutTorgbui Gorni Treve IV100% (2)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- AAA - 2010 - Diagnosis Treatment and Management of Children and Adults With Central Auditory Processing Disroder-AnnotatedDocument51 pagesAAA - 2010 - Diagnosis Treatment and Management of Children and Adults With Central Auditory Processing Disroder-AnnotatedadriricaldePas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Test Bank For Essentials of Dental Assisting 6th Edition by RobinsonDocument6 pagesTest Bank For Essentials of Dental Assisting 6th Edition by RobinsonRonald Ou100% (34)
- Restoration of Endodontically Treated TeethDocument7 pagesRestoration of Endodontically Treated TeethSuaeni Kurnia WirdaPas encore d'évaluation
- Picu Case StudyDocument3 pagesPicu Case Studyapi-455545776Pas encore d'évaluation
- Lichen PlanusDocument6 pagesLichen PlanusMerry MiraPas encore d'évaluation
- Dementia in Clinical Practice PDFDocument404 pagesDementia in Clinical Practice PDFadriricalde100% (1)
- Master Techniques in Orthopaedic Surgery Knee11Document1 139 pagesMaster Techniques in Orthopaedic Surgery Knee11hasla16Pas encore d'évaluation
- Newborn Physical ExaminationDocument17 pagesNewborn Physical ExaminationMaria Isabel de LeonPas encore d'évaluation
- Man Who Misttok His Wife For A HatDocument4 pagesMan Who Misttok His Wife For A HatOnyxKnightPas encore d'évaluation
- Fpsyg 07 01471Document15 pagesFpsyg 07 01471adriricaldePas encore d'évaluation
- Art:10.1007/s00455 016 9773 ZDocument12 pagesArt:10.1007/s00455 016 9773 ZadriricaldePas encore d'évaluation
- Myelomeningocele: How We Can Improve The Assessment of The Most Severe Form of Spina Bi FidaDocument7 pagesMyelomeningocele: How We Can Improve The Assessment of The Most Severe Form of Spina Bi FidaadriricaldePas encore d'évaluation
- Art:10 1007/BF00706692Document8 pagesArt:10 1007/BF00706692adriricaldePas encore d'évaluation
- English Et Al-2009-Developmental Disabilities Research ReviewsDocument7 pagesEnglish Et Al-2009-Developmental Disabilities Research ReviewsadriricaldePas encore d'évaluation
- Au Et Al-2010-Developmental Disabilities Research ReviewsDocument10 pagesAu Et Al-2010-Developmental Disabilities Research ReviewsadriricaldePas encore d'évaluation
- Tinnitus Current Understanding and Contemporary ManagementDocument6 pagesTinnitus Current Understanding and Contemporary ManagementadriricaldePas encore d'évaluation
- Velopharyngeal Insufficiency Diagnosis and ManagementDocument6 pagesVelopharyngeal Insufficiency Diagnosis and ManagementadriricaldePas encore d'évaluation
- Sellers Et Al-2014-Developmental Medicine & Child NeurologyDocument7 pagesSellers Et Al-2014-Developmental Medicine & Child NeurologyadriricaldePas encore d'évaluation
- 1 s2.0 S0030666516300469 MainDocument10 pages1 s2.0 S0030666516300469 MainadriricaldePas encore d'évaluation
- Structural Tympanic Membrane Changes in OMC and Cholesteatoma PDFDocument6 pagesStructural Tympanic Membrane Changes in OMC and Cholesteatoma PDFadriricaldePas encore d'évaluation
- Using The Language CharacteristicsDocument23 pagesUsing The Language CharacteristicsadriricaldePas encore d'évaluation
- Abnormal Language Pathway in Children With Angelman SyndromeDocument7 pagesAbnormal Language Pathway in Children With Angelman SyndromeadriricaldePas encore d'évaluation
- Cochlear Efferent Innervation and FunctionDocument7 pagesCochlear Efferent Innervation and FunctionadriricaldePas encore d'évaluation
- Bilateral Vestibulopathy PDFDocument6 pagesBilateral Vestibulopathy PDFadriricaldePas encore d'évaluation
- Delayed Developmental Language Milestones in Children With Duchenne's Muscular Dystrophy PDFDocument5 pagesDelayed Developmental Language Milestones in Children With Duchenne's Muscular Dystrophy PDFadriricaldePas encore d'évaluation
- Cochlear OtosclerosisDocument6 pagesCochlear OtosclerosisadriricaldePas encore d'évaluation
- Language and Eating Problems in Children PDFDocument10 pagesLanguage and Eating Problems in Children PDFadriricaldePas encore d'évaluation
- Auditory Development in Early Amplified ChildrenDocument20 pagesAuditory Development in Early Amplified ChildrenadriricaldePas encore d'évaluation
- Oromotor Variability in Children With Mild Pci PDFDocument10 pagesOromotor Variability in Children With Mild Pci PDFadriricaldePas encore d'évaluation
- Otolaryngology Head and Neck Surgery 2014 Dorresteijn 919 24Document6 pagesOtolaryngology Head and Neck Surgery 2014 Dorresteijn 919 24adriricaldePas encore d'évaluation
- Auris Nasus Larynx: Kimitaka KagaDocument11 pagesAuris Nasus Larynx: Kimitaka KagaadriricaldePas encore d'évaluation
- Feeding Disorders in ChildrenDocument16 pagesFeeding Disorders in ChildrenadriricaldePas encore d'évaluation
- Otolaryngology Head and Neck Surgery 2014 Wegner 201 7Document7 pagesOtolaryngology Head and Neck Surgery 2014 Wegner 201 7adriricaldePas encore d'évaluation
- Seminars in Fetal & Neonatal Medicine: Cristen Olds, John S. OghalaiDocument5 pagesSeminars in Fetal & Neonatal Medicine: Cristen Olds, John S. OghalaiadriricaldePas encore d'évaluation
- TalamoDocument7 pagesTalamoDanya RoblesPas encore d'évaluation
- Pain 3Document3 pagesPain 3Ka Yi MonicaPas encore d'évaluation
- 286 QsDocument5 pages286 Qsujangketul62Pas encore d'évaluation
- Cluster HeadacheDocument14 pagesCluster HeadacheadriantiariPas encore d'évaluation
- ExcerciseDocument7 pagesExcerciseprasadcshettyPas encore d'évaluation
- Arm CurlDocument3 pagesArm CurlNor AmalinaPas encore d'évaluation
- 2 KMCDocument39 pages2 KMCRafi MahendraPas encore d'évaluation
- A Project Proposal: Shri Jalaram Arogya Seva Trust HospitalDocument29 pagesA Project Proposal: Shri Jalaram Arogya Seva Trust HospitalSHREE SWAMINARAYAN AYURVEDIC COLLEGE GUJARATPas encore d'évaluation
- Abstracts Ecem13Document172 pagesAbstracts Ecem13opimadridistaPas encore d'évaluation
- And Neonatal Mechanical Ventilation. Berlin, Germany: Springer Verlag 2015Document3 pagesAnd Neonatal Mechanical Ventilation. Berlin, Germany: Springer Verlag 2015Ebrahem AlyPas encore d'évaluation
- Top College FeesDocument227 pagesTop College Feesarvindchopra3kPas encore d'évaluation
- Anatomy (Length: Duodenum 2.submucosaDocument12 pagesAnatomy (Length: Duodenum 2.submucosaURo KkuPas encore d'évaluation
- DaftarDocument2 pagesDaftarDini Dwi PuspitaPas encore d'évaluation
- Induction of Labor PDFDocument8 pagesInduction of Labor PDFGustomo PanantroPas encore d'évaluation
- Presion ArterialDocument133 pagesPresion ArterialAbelardo MorenoPas encore d'évaluation
- BSPD Clinical Holding Guidelines Final With Flow Chart 250416Document8 pagesBSPD Clinical Holding Guidelines Final With Flow Chart 250416druzair007Pas encore d'évaluation
- Usefulness of Presepsin (sCD14-ST) in Diagnosis of Infective Endocarditis - Preliminary Results of An Observational StudyDocument10 pagesUsefulness of Presepsin (sCD14-ST) in Diagnosis of Infective Endocarditis - Preliminary Results of An Observational StudyCristina GaidargiPas encore d'évaluation
- Policy On Pediatric Dental Pain ManagementDocument3 pagesPolicy On Pediatric Dental Pain ManagementمعتزباللهPas encore d'évaluation
- The Beacon - March 22, 2012Document18 pagesThe Beacon - March 22, 2012schaffnerpubPas encore d'évaluation
- Disruption of Radiologist WorkflowDocument6 pagesDisruption of Radiologist WorkflowChrisPas encore d'évaluation
- Tackla CVDocument3 pagesTackla CVapi-249693462Pas encore d'évaluation
- Determinants of Parental Decisions On 'Drop Out' From Cancer Treatment For Childhood Cancer PatientsDocument7 pagesDeterminants of Parental Decisions On 'Drop Out' From Cancer Treatment For Childhood Cancer PatientsAndrew HartonoPas encore d'évaluation
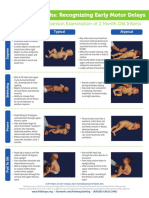
- 2 Months: Recognizing Early Motor Delays: A Comparison Examination of 2 Month Old InfantsDocument2 pages2 Months: Recognizing Early Motor Delays: A Comparison Examination of 2 Month Old InfantsMarinVaderŠamijaPas encore d'évaluation