Vous aimerez peut-être aussi
- Diet Rapidly Alters The Human Gut MicrobiomeDocument26 pagesDiet Rapidly Alters The Human Gut MicrobiomeGabriel CamarenaPas encore d'évaluation
- ElevatedatmosphericCO AltersrootsymbiontDocument12 pagesElevatedatmosphericCO AltersrootsymbiontGabriel CamarenaPas encore d'évaluation
- Climate Change Effects On ArbuscularDocument75 pagesClimate Change Effects On ArbuscularGabriel CamarenaPas encore d'évaluation
- The Diversity-Stability - McCann - NatureDocument6 pagesThe Diversity-Stability - McCann - Natureangstron328Pas encore d'évaluation
- Black Garlic - Alho NegroDocument9 pagesBlack Garlic - Alho NegroGustavo MartinsPas encore d'évaluation
- Effects of Marijuana Smoking On PulmonaryDocument8 pagesEffects of Marijuana Smoking On PulmonaryGabriel CamarenaPas encore d'évaluation
- Allium Thiosulfinates PDFDocument9 pagesAllium Thiosulfinates PDFGabriel CamarenaPas encore d'évaluation
- ElevatedatmosphericCO AltersrootsymbiontDocument12 pagesElevatedatmosphericCO AltersrootsymbiontGabriel CamarenaPas encore d'évaluation
- Climate Change E¡ects On B Ene Cial Plant MicroorganisminteractionsDocument18 pagesClimate Change E¡ects On B Ene Cial Plant MicroorganisminteractionsGabriel CamarenaPas encore d'évaluation
- Hormone QuestionnaireDocument8 pagesHormone QuestionnaireGabriel CamarenaPas encore d'évaluation
- Aged Garlic ExtractDocument17 pagesAged Garlic ExtractGabriel CamarenaPas encore d'évaluation
- Black Garlic ExtractDocument10 pagesBlack Garlic ExtractGabriel CamarenaPas encore d'évaluation
- Nutrition Implications For Fetal Alcohol PDFDocument18 pagesNutrition Implications For Fetal Alcohol PDFGabriel CamarenaPas encore d'évaluation
- Allium Thiosulfinates PDFDocument9 pagesAllium Thiosulfinates PDFGabriel CamarenaPas encore d'évaluation
- Hormone Cure Instructor Manual PDFDocument26 pagesHormone Cure Instructor Manual PDFGabriel Camarena83% (6)
- Oxidative Stress and Free Radicals in COPDDocument18 pagesOxidative Stress and Free Radicals in COPDGabriel CamarenaPas encore d'évaluation
- The Glucocorticoid Vulnerability HypothesisDocument20 pagesThe Glucocorticoid Vulnerability HypothesisGabriel CamarenaPas encore d'évaluation
- Open Access Definition of Inflammation, Causes of Inflammation PDFDocument9 pagesOpen Access Definition of Inflammation, Causes of Inflammation PDFGabriel CamarenaPas encore d'évaluation
- Subnutrition in The ElderlyDocument2 pagesSubnutrition in The ElderlyGabriel CamarenaPas encore d'évaluation
- Sensor Global de EstresDocument10 pagesSensor Global de EstresGabriel CamarenaPas encore d'évaluation
- Stress Exposure Food EmotionalDocument42 pagesStress Exposure Food EmotionalGabriel CamarenaPas encore d'évaluation
- The Song of The NeanderthalDocument1 pageThe Song of The NeanderthalGabriel CamarenaPas encore d'évaluation
- Stress Food InflammationDocument5 pagesStress Food InflammationGabriel CamarenaPas encore d'évaluation
- 2.1. Inflammation and Pro-Inflammatory MediatorsDocument23 pages2.1. Inflammation and Pro-Inflammatory MediatorsGabriel CamarenaPas encore d'évaluation
- Depression: The Nutrition Connection: Invited PapersDocument8 pagesDepression: The Nutrition Connection: Invited PapersGabriel CamarenaPas encore d'évaluation
- Eating Behavior and StressDocument8 pagesEating Behavior and StressGabriel CamarenaPas encore d'évaluation
- Depression: The Nutrition Connection: Invited PapersDocument8 pagesDepression: The Nutrition Connection: Invited PapersGabriel CamarenaPas encore d'évaluation
- "Manganese-Induced Neurotoxicity PDFDocument20 pages"Manganese-Induced Neurotoxicity PDFGabriel CamarenaPas encore d'évaluation
- Modification of Brain AgingDocument36 pagesModification of Brain AgingGabriel CamarenaPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5782)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (72)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- ASTM Collagen Standard PDFDocument14 pagesASTM Collagen Standard PDFAna RodriguezPas encore d'évaluation
- Avise, 2004. Molecular Markers, Natural History, and EvolutionDocument667 pagesAvise, 2004. Molecular Markers, Natural History, and EvolutionKaio BarrosPas encore d'évaluation
- Chemistry of TofuDocument6 pagesChemistry of TofuQuan ThànhPas encore d'évaluation
- Bryophytes and Heavy Metals: A Review Stanković D.J., Sabovljević D.A. I Sabovljević S.M., 2018.Document10 pagesBryophytes and Heavy Metals: A Review Stanković D.J., Sabovljević D.A. I Sabovljević S.M., 2018.AndrejaSKPas encore d'évaluation
- Protein Drug Binding Mechanisms and FactorsDocument22 pagesProtein Drug Binding Mechanisms and FactorsManikanta GupthaPas encore d'évaluation
- PyRosetta ManualDocument57 pagesPyRosetta Manualsisux9364Pas encore d'évaluation
- Applications of BioinformaticsDocument34 pagesApplications of BioinformaticsAntonMauntPas encore d'évaluation
- Sauces and Dressings A Review of Properties and ApplicationsDocument30 pagesSauces and Dressings A Review of Properties and ApplicationsJesus Rafael Zapata Alvarez100% (1)
- Membrane ProteinsDocument47 pagesMembrane ProteinsStanley ChikovePas encore d'évaluation
- Chimie Farma - Structure and Function of GPCRs (2019) PDFDocument299 pagesChimie Farma - Structure and Function of GPCRs (2019) PDFAlex-Bogdan VișaPas encore d'évaluation
- Human Nutriation DraftDocument66 pagesHuman Nutriation DraftFahadPas encore d'évaluation
- WRL0526Document45 pagesWRL0526NewdeersciPas encore d'évaluation
- Short Course On Basic Biology IDocument47 pagesShort Course On Basic Biology ICed HernandezPas encore d'évaluation
- Training Table To The Battlefield: Protein Recommendations For WarfightersDocument4 pagesTraining Table To The Battlefield: Protein Recommendations For WarfightersLalu Reza AldiraPas encore d'évaluation
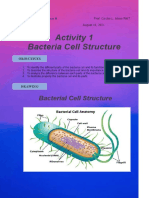
- Del Rosario Microbio Activity 1 Bacteria CellDocument6 pagesDel Rosario Microbio Activity 1 Bacteria CellNico LokoPas encore d'évaluation
- Pathogenesis of Dental Fluorosis: Biochemical and Cellular MechanismsDocument15 pagesPathogenesis of Dental Fluorosis: Biochemical and Cellular MechanismsAiluul AlmaaPas encore d'évaluation
- Whey Protein Based Edible Food Packaging Films and CoatingDocument25 pagesWhey Protein Based Edible Food Packaging Films and Coatingmnadia63Pas encore d'évaluation
- Qualitative color reactions identify protein componentsDocument3 pagesQualitative color reactions identify protein componentsPatricia ChongPas encore d'évaluation
- The Acupuncture System and The Liquid Crystalline Collagen Fibres of The Connective TissuesDocument17 pagesThe Acupuncture System and The Liquid Crystalline Collagen Fibres of The Connective Tissuesmkljhrguyt100% (2)
- Evoglow Basic Kit ManualDocument10 pagesEvoglow Basic Kit ManualPeter LeePas encore d'évaluation
- Chapter 4 ProteinDocument11 pagesChapter 4 ProteinAmbreen GhafoorPas encore d'évaluation
- Exam 2020Document34 pagesExam 2020Sameh NoorPas encore d'évaluation
- CV César ElenaDocument5 pagesCV César ElenajanePas encore d'évaluation
- Lipids, Biochem, ReviewerDocument9 pagesLipids, Biochem, ReviewerKerstine Anne AndradePas encore d'évaluation
- J Exp Med 1991 Linsley 721 30Document10 pagesJ Exp Med 1991 Linsley 721 30Fernanda RibeiroPas encore d'évaluation
- 1601 3863 1 PBDocument6 pages1601 3863 1 PBApriani GaPas encore d'évaluation
- Grade 10 STARS REVIEWERDocument12 pagesGrade 10 STARS REVIEWERjulianna startPas encore d'évaluation
- Five Overall Health-Improving Benefits of Royal JellyDocument2 pagesFive Overall Health-Improving Benefits of Royal JellyHayCurtis38Pas encore d'évaluation
- Reactions of Protein-01!11!2018Document26 pagesReactions of Protein-01!11!2018Rdh MnbPas encore d'évaluation
- Bio MoleculesDocument5 pagesBio MoleculesKashvi BachyasPas encore d'évaluation