Vous aimerez peut-être aussi
- Historia de La HistologíaDocument7 pagesHistoria de La HistologíaSilneleh F.Pas encore d'évaluation
- El Aparato CardiocirculatorioDocument18 pagesEl Aparato CardiocirculatorioJulio ReinaPas encore d'évaluation
- Cuestionario III.1 - CavidadesDocument19 pagesCuestionario III.1 - CavidadesPelea Con Pendejos TresmilPas encore d'évaluation
- Órganos de Los SentidosDocument3 pagesÓrganos de Los SentidosMarilynPas encore d'évaluation
- Origen Embriológico de RiñonDocument1 pageOrigen Embriológico de RiñonGlenn GarcíaPas encore d'évaluation
- Formulario de HistologiaDocument9 pagesFormulario de HistologiaShirleyLudyYajahuancaGaytanPas encore d'évaluation
- Anatomia Del Sistema Respiratorio Trabajo WordDocument10 pagesAnatomia Del Sistema Respiratorio Trabajo WordEduardoAriasPas encore d'évaluation
- Comunicación EndocrinaDocument1 pageComunicación EndocrinaJAIME GARRIDO VIVEROPas encore d'évaluation
- 4.°sec - Biologíanm - Ibim2020 - Ficha de Reforzamiento 1Document5 pages4.°sec - Biologíanm - Ibim2020 - Ficha de Reforzamiento 1Kiara BalcazarPas encore d'évaluation
- Capítulo 1. Introducción A La FarmacologíaDocument9 pagesCapítulo 1. Introducción A La FarmacologíaAlexandra ZaratePas encore d'évaluation
- Radiosensibilidad. Ley de Grotthus-Draper. Ley de Bergonié y Tribondeau. Radiolisis Del Agua. Bajas Dosis y Epidemiología.Document11 pagesRadiosensibilidad. Ley de Grotthus-Draper. Ley de Bergonié y Tribondeau. Radiolisis Del Agua. Bajas Dosis y Epidemiología.Angie RamírezPas encore d'évaluation
- 2222Document2 pages2222Jefferson Reyes M.Pas encore d'évaluation
- Problemas de Interpretacion HistológicaDocument4 pagesProblemas de Interpretacion HistológicaTaty Qs100% (1)
- Informe Del Corazon 02Document2 pagesInforme Del Corazon 02alvii saa100% (1)
- Cuestionario de Sistema Endocrino, HistologíaDocument10 pagesCuestionario de Sistema Endocrino, HistologíaDayana IntriagoPas encore d'évaluation
- CAPITULO 14 Comunicación de La Verdad, Intimidad, Confidencialidad y Secreto ProfesionalDocument11 pagesCAPITULO 14 Comunicación de La Verdad, Intimidad, Confidencialidad y Secreto Profesionaljoel menendezPas encore d'évaluation
- Hueso AtlasDocument6 pagesHueso AtlasHugo GonzalezPas encore d'évaluation
- Informe Practica Inmovilizacion SuperiorDocument5 pagesInforme Practica Inmovilizacion SuperiorErick EstévezPas encore d'évaluation
- Histología de Los Ganglios LinfáticosDocument10 pagesHistología de Los Ganglios LinfáticosDiana Marcela AcostaPas encore d'évaluation
- Sistema DigestivoDocument55 pagesSistema DigestivoPamelaPas encore d'évaluation
- Actividad 3. Órganos Primarios y Secundarios Del Sistema InmuneDocument10 pagesActividad 3. Órganos Primarios y Secundarios Del Sistema InmuneSarai GomezPas encore d'évaluation
- Informe de Generalidades de La Genetica y Protocolos de BioseguridadDocument8 pagesInforme de Generalidades de La Genetica y Protocolos de BioseguridadRosali Sara Ccuno QuispePas encore d'évaluation
- Histologia de LaringeDocument3 pagesHistologia de LaringeAdnan Haj Yousef Cordero100% (1)
- Histología de Los Vasos SanguíneosDocument13 pagesHistología de Los Vasos SanguíneosGeorgina GalindoPas encore d'évaluation
- BarrerasDocument3 pagesBarrerasAbraham guzman MateoPas encore d'évaluation
- Introduccion Embriología Del Sistema RespiratorioDocument1 pageIntroduccion Embriología Del Sistema Respiratoriojorge100% (1)
- Vía Olfatoria y Gustativa TerminadoDocument8 pagesVía Olfatoria y Gustativa Terminadobastian carreñoPas encore d'évaluation
- Mapa Conceptual HistologiaDocument3 pagesMapa Conceptual Histologialisbeth corderoPas encore d'évaluation
- Histología de La Cavidad NasalDocument2 pagesHistología de La Cavidad NasalGiovanni Saúl Contla Trejo86% (7)
- Diapositivas ExposiciónDocument33 pagesDiapositivas ExposiciónNataly PillajoPas encore d'évaluation
- Informe N8Document6 pagesInforme N8Solange YánezPas encore d'évaluation
- Esquema Comparativo Sobre La Contracción El Músculo Cardiaco y El Músculo EsqueléticoDocument2 pagesEsquema Comparativo Sobre La Contracción El Músculo Cardiaco y El Músculo EsqueléticoJulia EsquivelPas encore d'évaluation
- La Sustancia BlancaDocument2 pagesLa Sustancia BlancaDamaris Yólatl Martínez JiménezPas encore d'évaluation
- Histología Del Aparato Respiratorio InferiorDocument24 pagesHistología Del Aparato Respiratorio InferiorJeanett Velásquez CorralesPas encore d'évaluation
- Mapa Mental Sistema CirculatorioDocument1 pageMapa Mental Sistema CirculatorioValentina PedrazaPas encore d'évaluation
- Sesion 48 Enfermedades Infecciosas Del Aparato DigestivoDocument29 pagesSesion 48 Enfermedades Infecciosas Del Aparato DigestivoPacko Guerrero CastroPas encore d'évaluation
- Ta 1 Épuli Fribromatoso en EquinosDocument22 pagesTa 1 Épuli Fribromatoso en EquinosEdwin Alvarado MuñozPas encore d'évaluation
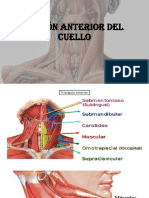
- Región Anterior Del CuelloDocument14 pagesRegión Anterior Del CuelloBillyPas encore d'évaluation
- Practica 3 y 4 Diversidad CelularDocument4 pagesPractica 3 y 4 Diversidad CelularmarulasPas encore d'évaluation
- Fisiologia Regulaciondelarespiracion 140916092221 Phpapp01Document2 pagesFisiologia Regulaciondelarespiracion 140916092221 Phpapp01Fabiola SanchezPas encore d'évaluation
- Anato II Ciclo IIDocument21 pagesAnato II Ciclo IIDiegoPas encore d'évaluation
- Olfato y Gusto ExpDocument21 pagesOlfato y Gusto ExpKEYLA DENISS RAMIREZ DIAZPas encore d'évaluation
- Tema 12. Intestino Anterior Medio y PosteriorDocument10 pagesTema 12. Intestino Anterior Medio y PosteriorVibe Nation100% (1)
- Funcion de Alantoides Saco VitelinoDocument1 pageFuncion de Alantoides Saco VitelinovioletaPas encore d'évaluation
- Medicina PrehistóricaDocument2 pagesMedicina PrehistóricaACPas encore d'évaluation
- Sistema CirculatorioDocument15 pagesSistema CirculatorioElvismar Perez100% (2)
- Las ArteriasDocument6 pagesLas ArteriasMarcos Erli Banda CubasPas encore d'évaluation
- Procesamiento de Linfocitos y ClonesDocument5 pagesProcesamiento de Linfocitos y ClonesKenneth MazacónPas encore d'évaluation
- Universidad Tecnológica de Santiag1Document4 pagesUniversidad Tecnológica de Santiag1Henrius FeguensPas encore d'évaluation
- Practica 4. Epitelios de RevestimientoDocument21 pagesPractica 4. Epitelios de RevestimientoNorma GLPas encore d'évaluation
- Principios Generales de La Funcion GastrointestinalDocument34 pagesPrincipios Generales de La Funcion GastrointestinalOscar AgüeroPas encore d'évaluation
- Neurofisiología Motora e IntegradoraDocument20 pagesNeurofisiología Motora e IntegradorazafraPas encore d'évaluation
- 25 Palabras Conforme El Abecedario Del Sistema Digestivo Hasta Historia Del BaloncestoDocument5 pages25 Palabras Conforme El Abecedario Del Sistema Digestivo Hasta Historia Del BaloncestoFrheddyyRuizPas encore d'évaluation
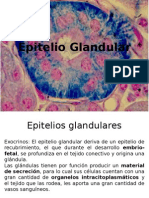
- Epitelio GlandularDocument49 pagesEpitelio GlandularSebastian Mariscal BarraPas encore d'évaluation
- Alteraciones en El Desarrollo EmbrionarioDocument7 pagesAlteraciones en El Desarrollo EmbrionarioLaura DiazPas encore d'évaluation
- AlveolosDocument10 pagesAlveolosAlex RomeroPas encore d'évaluation
- Las Características Histológicas de Las Vías Urinarias Excretoras Son SimilaresDocument3 pagesLas Características Histológicas de Las Vías Urinarias Excretoras Son Similaresmaalhues100% (1)
- PPT# 1 Anatomia de La MamaDocument23 pagesPPT# 1 Anatomia de La MamaAbel Garcia smithPas encore d'évaluation
- Antología Bioquímica Q.F.B. Melba Fernandez RojasDocument42 pagesAntología Bioquímica Q.F.B. Melba Fernandez RojasEdgarmarquezPas encore d'évaluation
- Biografias MedicinaDocument11 pagesBiografias MedicinaAlanPas encore d'évaluation
- Taller de Medidas en Epidemiologia Número 2Document3 pagesTaller de Medidas en Epidemiologia Número 2jamrPas encore d'évaluation
- Micosis Sistemicas Infex 2Document29 pagesMicosis Sistemicas Infex 2jamrPas encore d'évaluation
- Taller de Medidas en Epidemiologia Número 2Document3 pagesTaller de Medidas en Epidemiologia Número 2jamrPas encore d'évaluation
- Desarrollo Del Feto y Estructura de La PlacentaDocument11 pagesDesarrollo Del Feto y Estructura de La PlacentajamrPas encore d'évaluation
- Enfermedad de Pompe. Bioquimica 1Document4 pagesEnfermedad de Pompe. Bioquimica 1jamrPas encore d'évaluation
- Empirismo, Sensacionalismo y Positivismo. TallerDocument6 pagesEmpirismo, Sensacionalismo y Positivismo. Tallerjamr0% (1)
- Tema 8 Celula IIDocument8 pagesTema 8 Celula IIRocio CavieresPas encore d'évaluation
- Los GranulocitosDocument2 pagesLos GranulocitosRafael Aristides AcostaPas encore d'évaluation
- Respiración CelularDocument16 pagesRespiración CelularAlvion LozcaPas encore d'évaluation
- Tejido ConjuntivoDocument20 pagesTejido ConjuntivoTiciana GarciaPas encore d'évaluation
- CUESTIONARIO Unidad 1Document12 pagesCUESTIONARIO Unidad 1muriel aravena makucPas encore d'évaluation
- Catalina Cava Tesis DoctoralDocument231 pagesCatalina Cava Tesis DoctoralRoberto Morales RiveraPas encore d'évaluation
- Histologia Del HigadoDocument9 pagesHistologia Del HigadoErich MaicolPas encore d'évaluation
- Guia de Aprendizaje AutonomoDocument2 pagesGuia de Aprendizaje Autonomosaxtenor6Pas encore d'évaluation
- Extración Del Adn CromosomalDocument4 pagesExtración Del Adn CromosomalRafael Soria GalvarroPas encore d'évaluation
- Generar ResultadoDocument3 pagesGenerar ResultadovivivanaPas encore d'évaluation
- Micro Biolog I ADocument42 pagesMicro Biolog I AGianpierre Jhonatan ChavezPas encore d'évaluation
- Cuardenillo Pato 2024Document395 pagesCuardenillo Pato 2024Luiz FelipePas encore d'évaluation
- Examen de Biologïa Segundo MedioDocument14 pagesExamen de Biologïa Segundo Mediouzielx100% (1)
- Biologia 08 - Resumen Orne 2do ParcialDocument13 pagesBiologia 08 - Resumen Orne 2do ParcialSantino CerezoPas encore d'évaluation
- R6 E5 Actividad-MetabolicaDocument6 pagesR6 E5 Actividad-MetabolicaGómez AbrilPas encore d'évaluation
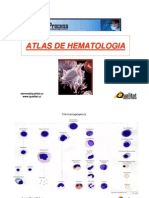
- Atlas HematologiaDocument83 pagesAtlas HematologiaGamaliel Canté HdezPas encore d'évaluation
- Fotosíntesis Respiración CelularDocument3 pagesFotosíntesis Respiración CelularTumamita3000Pas encore d'évaluation
- LA CÉLULA y LOS VIRUSDocument44 pagesLA CÉLULA y LOS VIRUSmarcelaPas encore d'évaluation
- Articulo Cientifico Raiz de CebollaDocument8 pagesArticulo Cientifico Raiz de CebollaMaria Camila MedinaPas encore d'évaluation
- Celula Aucariota - CitoplasmaDocument5 pagesCelula Aucariota - CitoplasmaDenner Kilder Cabrejos RamirezPas encore d'évaluation
- 1 UNIDAD La Nutricion y La Celula Grado 4 (Document14 pages1 UNIDAD La Nutricion y La Celula Grado 4 (chela solano rodriguezPas encore d'évaluation
- INFORME DE LABORATORIO - Tinción de Gram.Document6 pagesINFORME DE LABORATORIO - Tinción de Gram.Angelee Pacheco Rubio50% (2)
- ParaclinicosDocument2 pagesParaclinicosYuliana VanegasPas encore d'évaluation
- Solución Taller N°1 Ácidos NucléicosDocument10 pagesSolución Taller N°1 Ácidos NucléicosJuly Ramírez100% (2)
- Bme U3 A2 VesrDocument5 pagesBme U3 A2 VesrMonse SanchezPas encore d'évaluation
- Introduccion Anatomia VegetalDocument70 pagesIntroduccion Anatomia VegetalAnonymous iWhxcenPas encore d'évaluation
- Actividad 4Document4 pagesActividad 4Mayra Graciela Miranda SuarezPas encore d'évaluation
- UNT SILABO B.M.y C. 2023Document13 pagesUNT SILABO B.M.y C. 2023BRAYAN JOEL ANDRADE OLIVARESPas encore d'évaluation
- Célula ProcariotaDocument11 pagesCélula ProcariotaHazael Bautista RamírezPas encore d'évaluation
- Instituto Politécnico Nacional. Escuela Nacional de Medicina y HomeopatíaDocument29 pagesInstituto Politécnico Nacional. Escuela Nacional de Medicina y HomeopatíaHernández Ortega YolotzinPas encore d'évaluation