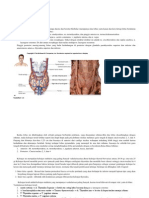
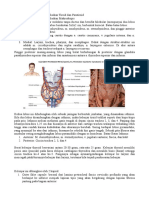
Vous aimerez peut-être aussi
- Drug Induced HepatitisDocument30 pagesDrug Induced HepatitisRamadhan Ananda Putra100% (2)
- Sawar Darah OtakDocument9 pagesSawar Darah OtakTibo ColdPas encore d'évaluation
- Laporan Kasus NefrolitiasisDocument10 pagesLaporan Kasus NefrolitiasisRamadhan Ananda Putra100% (1)
- Disfungsi EndotelDocument13 pagesDisfungsi EndotelAngga HadityaPas encore d'évaluation
- Tinjauan PustakaDocument12 pagesTinjauan Pustakasemarang 21Pas encore d'évaluation
- SSc-ILD Lengkap NEW - 26012021Document27 pagesSSc-ILD Lengkap NEW - 26012021Dewi AstariniPas encore d'évaluation
- PBL Endokrin Skenario 2Document23 pagesPBL Endokrin Skenario 2Ariandhy EchoPas encore d'évaluation
- ANGIOGENESIS Patofisiologi Dan AplikasiDocument36 pagesANGIOGENESIS Patofisiologi Dan AplikasiIftinan LQPas encore d'évaluation
- Life-11-00986 (1) en IdDocument16 pagesLife-11-00986 (1) en IdfebrynnadaPas encore d'évaluation
- UAS Assoc. Prof. DR - Dr. H Agung Putra, M.si. Med - Rheza Rizaldy - MBK2016010210Document8 pagesUAS Assoc. Prof. DR - Dr. H Agung Putra, M.si. Med - Rheza Rizaldy - MBK2016010210Rheza RizaldyPas encore d'évaluation
- HeparinDocument16 pagesHeparinUploader AlbumPas encore d'évaluation
- 817 3295 1 PB PDFDocument12 pages817 3295 1 PB PDFrisnaPas encore d'évaluation
- Fisiologi Koroner AterosklerosisDocument15 pagesFisiologi Koroner AterosklerosisjarotPas encore d'évaluation
- 1610 1610 1283 Referat Disfungsi EndotelDocument26 pages1610 1610 1283 Referat Disfungsi Endotelolivia magdalenaPas encore d'évaluation
- HormonDocument25 pagesHormonGolden GamingPas encore d'évaluation
- Makalah Perkembangan HewanDocument12 pagesMakalah Perkembangan HewanPratiwiPas encore d'évaluation
- Tugas Pendahuluan - Fisiologi Sumsum TulangDocument15 pagesTugas Pendahuluan - Fisiologi Sumsum TulangKielgrey K'Pas encore d'évaluation
- AbstrakDocument15 pagesAbstrakKrisna FernandaPas encore d'évaluation
- Tugas Hematologi GegeDocument17 pagesTugas Hematologi GegeAngeline LoisyePas encore d'évaluation
- Sistem Imun & Peredaran Darah - Acara IiDocument13 pagesSistem Imun & Peredaran Darah - Acara IiBima Surya PratamaPas encore d'évaluation
- ANGIOGENESIS Patofisiologi Dan AplikasiDocument51 pagesANGIOGENESIS Patofisiologi Dan AplikasiIftinan LQ100% (1)
- Skenario 2 GondokDocument29 pagesSkenario 2 GondokseshabrinaPas encore d'évaluation
- Sistym EndokrinDocument25 pagesSistym Endokrinputihkertas427Pas encore d'évaluation
- Her BioselDocument4 pagesHer BioselDefli Yuandika RPas encore d'évaluation
- SEL ENDOTEL (DR - DR - .Chodidjah, M.Kes)Document22 pagesSEL ENDOTEL (DR - DR - .Chodidjah, M.Kes)Rakhmat RamadhaniPas encore d'évaluation
- Preskas-Dr Enos-Karsinoma Tiroid-IlyasDocument45 pagesPreskas-Dr Enos-Karsinoma Tiroid-IlyasJamaluddinLukmanPas encore d'évaluation
- Inti ProteinDocument3 pagesInti Proteinnugraha123Pas encore d'évaluation
- HemopoiesisDocument7 pagesHemopoiesisJean StepaniPas encore d'évaluation
- Sistem HematologiDocument24 pagesSistem HematologiHandis WitantokoPas encore d'évaluation
- LP Nodul TiroidDocument30 pagesLP Nodul TiroidEZ Syakal100% (3)
- S2 2015 337216 Chapter5Document25 pagesS2 2015 337216 Chapter5dini hanifaPas encore d'évaluation
- Makalah Ske 6 FixDocument28 pagesMakalah Ske 6 Fixazam ahmedPas encore d'évaluation
- Fisiologi Sistem Endokrin Ikan TunaDocument9 pagesFisiologi Sistem Endokrin Ikan TunaNur Roudlotul LailaPas encore d'évaluation
- Sari Pustaka I Perisit RevisiDocument27 pagesSari Pustaka I Perisit Revisidrrizqiwahyu85Pas encore d'évaluation
- Tugas Pembelahan SelDocument8 pagesTugas Pembelahan SelOsi Arosi OchiePas encore d'évaluation
- AstrocytomaDocument24 pagesAstrocytomaFira AzisPas encore d'évaluation
- Mekanisme Apoptosis Pada SepsisDocument9 pagesMekanisme Apoptosis Pada SepsisSurya TriwijayaPas encore d'évaluation
- Hitung LeukositDocument10 pagesHitung LeukositNayspisces Nyea NoelupthflenttheaPas encore d'évaluation
- PhocomeliaDocument6 pagesPhocomeliafaris nizarPas encore d'évaluation
- Bab 2Document22 pagesBab 2Safril Ilmi RamadhanPas encore d'évaluation
- PBL Blok 11Document5 pagesPBL Blok 11jessicaPas encore d'évaluation
- STT Tumor - ObbiqDocument51 pagesSTT Tumor - Obbiqnnda143Pas encore d'évaluation
- Jurnal Inflamasi TbaDocument14 pagesJurnal Inflamasi TbaFitri Andriani IbrahimPas encore d'évaluation
- Patofisiologi-Polymyalgia RheumaticaDocument2 pagesPatofisiologi-Polymyalgia RheumaticafuadanharuddinfunzhuPas encore d'évaluation
- TMSK2 0111 Putri Kurnia AssiqyDocument21 pagesTMSK2 0111 Putri Kurnia AssiqyPutri Kurnia AssiqyPas encore d'évaluation
- Materi Biomedik IiDocument95 pagesMateri Biomedik Iiadan erPas encore d'évaluation
- Review Jurnal KeduaDocument4 pagesReview Jurnal Keduanaufal umerPas encore d'évaluation
- 12 Desember 2013 - HPS TKV Bab VascularDocument108 pages12 Desember 2013 - HPS TKV Bab VascularJuwita Rashi LumbantoruanPas encore d'évaluation
- Lapsus Tumor Tiroid BaiqDocument30 pagesLapsus Tumor Tiroid BaiqihwanuPas encore d'évaluation
- Tugas Baca Maria A.Y. FernandezDocument19 pagesTugas Baca Maria A.Y. FernandezAnonymous hYJRYhXPas encore d'évaluation
- Siklus SelDocument20 pagesSiklus SelVante TaniePas encore d'évaluation
- ANGIOGENESIS Patofisiologi Dan AplikasiDocument16 pagesANGIOGENESIS Patofisiologi Dan AplikasiAriqa AzzahraPas encore d'évaluation
- Soft Tissue TumorDocument48 pagesSoft Tissue TumorMustafa HolidiPas encore d'évaluation
- ThyroidDocument29 pagesThyroidIr Ardy YusriawanPas encore d'évaluation
- Referat Nodul TiroidDocument19 pagesReferat Nodul TiroidbundamegaPas encore d'évaluation
- Referat - Penatalaksanaan Thyroid NoduleDocument29 pagesReferat - Penatalaksanaan Thyroid NoduleSilvia ValentinaPas encore d'évaluation
- Sel Darah PDFDocument39 pagesSel Darah PDFIrvi Firqotul Aini87% (15)
- Vaskular BookDocument151 pagesVaskular BookJuwita Rashi Lumbantoruan100% (1)
- Dokumen - Tips Laporan Kasus SGBDocument20 pagesDokumen - Tips Laporan Kasus SGBScribdMeUpPas encore d'évaluation
- Portofolio KDK+BPDocument7 pagesPortofolio KDK+BPRamadhan Ananda PutraPas encore d'évaluation
- Case Vesikolithiasis AnakDocument24 pagesCase Vesikolithiasis AnakRamadhan Ananda PutraPas encore d'évaluation
- Instrumen PuskesmasDocument124 pagesInstrumen PuskesmasRamadhan Ananda Putra100% (12)
- Kejang DemamDocument1 pageKejang DemamAstri Ayu SetianingtyasPas encore d'évaluation
- Portofolio BPHDocument18 pagesPortofolio BPHRamadhan Ananda PutraPas encore d'évaluation
- Gambaran Radiologi HepatomaDocument33 pagesGambaran Radiologi HepatomaRamadhan Ananda PutraPas encore d'évaluation
- Laporan Kasus HipospadiaDocument23 pagesLaporan Kasus HipospadiaAdi RiyadliPas encore d'évaluation
- Laporan Kasus KolestasisDocument4 pagesLaporan Kasus KolestasisRamadhan Ananda PutraPas encore d'évaluation
- Laporan Kasus Vesikolithiasis AnakDocument24 pagesLaporan Kasus Vesikolithiasis AnakRamadhan Ananda PutraPas encore d'évaluation
- Referat Radio TB CoxitisDocument31 pagesReferat Radio TB CoxitisRamadhan Ananda PutraPas encore d'évaluation
- Laporan Kasus ApendisitisDocument17 pagesLaporan Kasus ApendisitisRamadhan Ananda PutraPas encore d'évaluation
- Kolestasis EkstrahepatalDocument5 pagesKolestasis EkstrahepatalRamadhan Ananda PutraPas encore d'évaluation
- Referat Radiologi BPH Dan Kanker ProstatDocument34 pagesReferat Radiologi BPH Dan Kanker ProstatRamadhan Ananda PutraPas encore d'évaluation
- PoaDocument46 pagesPoaRamadhan Ananda PutraPas encore d'évaluation
- Analisis Kasus TBDocument4 pagesAnalisis Kasus TBRamadhan Ananda PutraPas encore d'évaluation