Vous aimerez peut-être aussi
- Skin Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsD'EverandSkin Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsPas encore d'évaluation
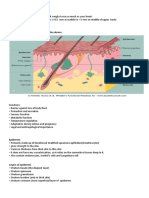
- Anatomy and Physiology of The Skin.3Document11 pagesAnatomy and Physiology of The Skin.3alinaPas encore d'évaluation
- Skin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsD'EverandSkin cell, Functions, Diseases, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsPas encore d'évaluation
- Lecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalDocument24 pagesLecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalAli Waheed jolan100% (1)
- Anatomy of The Skin: Ectodermal Tissue Muscles Bones Ligaments Internal OrganDocument4 pagesAnatomy of The Skin: Ectodermal Tissue Muscles Bones Ligaments Internal OrganKaizen Science Academy100% (1)
- The Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemD'EverandThe Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemPas encore d'évaluation
- Administer An Intradermal InjectionDocument3 pagesAdminister An Intradermal InjectionGurinder GillPas encore d'évaluation
- Skin Pigmentation + Hari DisorderDocument113 pagesSkin Pigmentation + Hari DisorderAfiqah So JasmiPas encore d'évaluation
- Skin Diseases Affecting The VulvaDocument7 pagesSkin Diseases Affecting The VulvaNoraPas encore d'évaluation
- Skin DisordersDocument202 pagesSkin DisordersMj Briones100% (1)
- Function of The SkinDocument69 pagesFunction of The SkinapermatagamaPas encore d'évaluation
- Plasma Fibroblast TherapyDocument24 pagesPlasma Fibroblast TherapyDrAmit Gaba MdsPas encore d'évaluation
- Sun Protection in Acne, Dr. Dwi Retno Adi WinarniDocument29 pagesSun Protection in Acne, Dr. Dwi Retno Adi WinarniAdi Sutriwanto PasaribuPas encore d'évaluation
- Skin Disease WartsDocument4 pagesSkin Disease WartsFatima Al MarzouqiPas encore d'évaluation
- Language of DermatologyDocument45 pagesLanguage of DermatologykurutalaPas encore d'évaluation
- Introduction To Dermatology Assessment of A Dermatologic PatientDocument86 pagesIntroduction To Dermatology Assessment of A Dermatologic PatientAtifPas encore d'évaluation
- Pigmentary Disorders: Mikias Woldetensay Dermatology & Venereology OCT 2021Document40 pagesPigmentary Disorders: Mikias Woldetensay Dermatology & Venereology OCT 2021CHALIE MEQU100% (1)
- Aplied Anatomy BotoxDocument8 pagesAplied Anatomy BotoxYesuratnam DudduPas encore d'évaluation
- The History of Botulinum Toxin From Poison To BeautyDocument3 pagesThe History of Botulinum Toxin From Poison To BeautyElaine MedeirosPas encore d'évaluation
- AcneDocument13 pagesAcneOktiya SariPas encore d'évaluation
- Superficial Fungal Infections of The Skin VoiceoverDocument67 pagesSuperficial Fungal Infections of The Skin VoiceoverMedo JT100% (1)
- Dermatology FinalDocument16 pagesDermatology FinalMelinda100% (2)
- Principles of Topical Treatments in DermatologyDocument18 pagesPrinciples of Topical Treatments in DermatologyAnonymous OlS0WZwPas encore d'évaluation
- Anatomy&Physiology of SkinDocument45 pagesAnatomy&Physiology of Skinpreet kaurPas encore d'évaluation
- ReviewerDocument15 pagesReviewerYza Grace S. PayangdoPas encore d'évaluation
- SkinDocument38 pagesSkinrodelagapito100% (1)
- Human Skin 1Document9 pagesHuman Skin 1Baciu DianaPas encore d'évaluation
- An Approach To The Patient With ErythrodermaDocument42 pagesAn Approach To The Patient With ErythrodermaShakilPas encore d'évaluation
- Srinadh Neonatal Skin Conditions FinalDocument110 pagesSrinadh Neonatal Skin Conditions FinalSrinadh Pragada100% (1)
- Assalamu'alikum WR WB: Harijono KariosentonoDocument59 pagesAssalamu'alikum WR WB: Harijono KariosentonoDhiastikaPas encore d'évaluation
- Dermatology PDFDocument41 pagesDermatology PDFmariatanPas encore d'évaluation
- RetinoidDocument22 pagesRetinoidPresstisa AxeliaPas encore d'évaluation
- Skin Rejuvenation Treatments PDFDocument7 pagesSkin Rejuvenation Treatments PDFNony Satya Anugrah100% (1)
- Neuromodulators Injection On The ForeheadDocument24 pagesNeuromodulators Injection On The ForeheadJennifer GomezPas encore d'évaluation
- Skin Anatomy and PhysiologyDocument5 pagesSkin Anatomy and PhysiologyKhan KamaalPas encore d'évaluation
- Skin and Its AppendagesDocument28 pagesSkin and Its AppendagesTK Balbin Liquit100% (1)
- Jett Plasma Lift Medical Manual ENG - Akt - .9.9.15 PDFDocument27 pagesJett Plasma Lift Medical Manual ENG - Akt - .9.9.15 PDFMichaely Natali100% (1)
- Skin PDFDocument57 pagesSkin PDFravindra sharma100% (1)
- Mesotherapy Poster PDFDocument2 pagesMesotherapy Poster PDFgemm88Pas encore d'évaluation
- Serdev Suture Techniques Breast Lift - Prof Nikolay P Serdev MD PHDDocument4 pagesSerdev Suture Techniques Breast Lift - Prof Nikolay P Serdev MD PHDjobetobaPas encore d'évaluation
- "Vision Without Action Is A Daydream. Action Without Vision Is A Nightmare." Japanese ProverbDocument22 pages"Vision Without Action Is A Daydream. Action Without Vision Is A Nightmare." Japanese Proverb123456100% (1)
- Dermapen Cryo Users ManualDocument44 pagesDermapen Cryo Users ManualDIANAPas encore d'évaluation
- Scars PDFDocument9 pagesScars PDFMichaely NataliPas encore d'évaluation
- Structural Fat GraftingDocument19 pagesStructural Fat GraftingGautam KalraPas encore d'évaluation
- Aesthetic Medicine - Part 2Document9 pagesAesthetic Medicine - Part 2unknownPas encore d'évaluation
- DermatitisDocument6 pagesDermatitisNader SmadiPas encore d'évaluation
- Skin Structure and FunctionsDocument31 pagesSkin Structure and FunctionsAfifa Prima Gitta100% (2)
- 05-BoNtA 568Document44 pages05-BoNtA 568Yovie Prayekti100% (2)
- Infraorbital Dark Circles - A Review of The Pathogenesis, Evaluation and TreatmentDocument8 pagesInfraorbital Dark Circles - A Review of The Pathogenesis, Evaluation and TreatmentElaine MedeirosPas encore d'évaluation
- Sec 4 - Techniques 2Document31 pagesSec 4 - Techniques 2Karem TubePas encore d'évaluation
- Acne Vulgaris (Or Acne) Is A CommonDocument21 pagesAcne Vulgaris (Or Acne) Is A CommonNia YuniarPas encore d'évaluation
- See Full Prescribing Information For Complete Boxed WarningDocument22 pagesSee Full Prescribing Information For Complete Boxed WarningTracy PopePas encore d'évaluation
- Integumentary: Skin, Hair, Nails, GlandsDocument67 pagesIntegumentary: Skin, Hair, Nails, GlandsJosephPas encore d'évaluation
- Skin Hydration and Hyaluronic AcidDocument5 pagesSkin Hydration and Hyaluronic AcidChris GohPas encore d'évaluation
- 7 Bleeding Disorders DR - AlaaDocument35 pages7 Bleeding Disorders DR - Alaaaj1998ajPas encore d'évaluation
- Anatomi Fisiologi KulitDocument80 pagesAnatomi Fisiologi Kulitedo100% (1)
- PRS 2015 01 SykesDocument15 pagesPRS 2015 01 Sykespsdsportsdoc100% (2)
- Actinic Keratosis HandoutDocument1 pageActinic Keratosis HandoutNicco Jake AlegroPas encore d'évaluation
- Poly L Lactic Acid SicklesDocument4 pagesPoly L Lactic Acid SicklesRafael Autran Cavalcante AraújoPas encore d'évaluation
- PHS 205 Cardiovascular SystemDocument33 pagesPHS 205 Cardiovascular Systemdivineraymond34Pas encore d'évaluation
- Puberphonia 11Document19 pagesPuberphonia 11jaguar1979100% (2)
- Seminar On Factor Affecting Drug Absorption: Presented byDocument42 pagesSeminar On Factor Affecting Drug Absorption: Presented byKrunal SamejaPas encore d'évaluation
- BCA Protocol PDFDocument7 pagesBCA Protocol PDFAlyssa236Pas encore d'évaluation
- Allergy Genu Ection? It's Surmount With Special Focus On Ear, Nose and ThroatDocument11 pagesAllergy Genu Ection? It's Surmount With Special Focus On Ear, Nose and ThroatAna BrankovićPas encore d'évaluation
- HURST NCLEX Review Facts PDFDocument6 pagesHURST NCLEX Review Facts PDFMikePas encore d'évaluation
- Biochemistry Quizzes by Ronnie BaticulonDocument20 pagesBiochemistry Quizzes by Ronnie BaticulonMhartin GarciaPas encore d'évaluation
- Reactive The Trauma Active Brain WorksheetDocument5 pagesReactive The Trauma Active Brain Worksheetkadbury1900Pas encore d'évaluation
- Session 1 - Electrolyte and Degree of Dissociation - 2022Document39 pagesSession 1 - Electrolyte and Degree of Dissociation - 2022Anjal MallickPas encore d'évaluation
- General Principles of Exercise For Health and Fitness: © 2014 Pearson Education, IncDocument24 pagesGeneral Principles of Exercise For Health and Fitness: © 2014 Pearson Education, IncmichellePas encore d'évaluation
- Bad Blood CardsDocument15 pagesBad Blood CardsEvry BoyTestPas encore d'évaluation
- Liceo de Cagayan University: Acute AppendicitisDocument2 pagesLiceo de Cagayan University: Acute AppendicitisKylie AstrudPas encore d'évaluation
- CRANIECTOMY OR RotationDocument41 pagesCRANIECTOMY OR RotationFretzgine Lou ManuelPas encore d'évaluation
- Cholesterol SynthesisDocument19 pagesCholesterol Synthesisbrian mgabiPas encore d'évaluation
- Structure of The Lactating Breast: Lactation Is The Process by Which Milk Is Synthesized and Secreted From The MammaryDocument5 pagesStructure of The Lactating Breast: Lactation Is The Process by Which Milk Is Synthesized and Secreted From The Mammaryratna putriPas encore d'évaluation
- Exudate ManagementDocument6 pagesExudate Managementmsaunit1Pas encore d'évaluation
- Plant Agronomic Performance and Selection For Green Roofs Under Mediterranean and Maritime Climate ConditionsDocument160 pagesPlant Agronomic Performance and Selection For Green Roofs Under Mediterranean and Maritime Climate ConditionsPoly LitoikairoiPas encore d'évaluation
- Thinking Like A Plant - HoldregeDocument4 pagesThinking Like A Plant - HoldregeMartin BungePas encore d'évaluation
- Silva Meditation 5 Fall Asleep - Alarm - Stay AwakeDocument4 pagesSilva Meditation 5 Fall Asleep - Alarm - Stay AwakeMichiel PaulusPas encore d'évaluation
- Cardiovascular Blood VesselsDocument12 pagesCardiovascular Blood VesselswatuwaitingforPas encore d'évaluation
- 10 Current Research Articles Related To Our SystemDocument11 pages10 Current Research Articles Related To Our Systemhomework solutionPas encore d'évaluation
- (Equine Veterinary Journal 2010-Jan 05 Vol. 37 Iss. 2) W. W. MUIR - Pain Therapy in Horses (2010) (10.2746 - 0425164054223831) - Libgen - LiDocument3 pages(Equine Veterinary Journal 2010-Jan 05 Vol. 37 Iss. 2) W. W. MUIR - Pain Therapy in Horses (2010) (10.2746 - 0425164054223831) - Libgen - LiAcupuntura de EquinosPas encore d'évaluation
- Acute Coronary Syndrome, STEMI, Anterior Wall, Killips - 1, DM Type II - UncontrolledDocument61 pagesAcute Coronary Syndrome, STEMI, Anterior Wall, Killips - 1, DM Type II - UncontrolledJai - Ho100% (5)
- Cardiovascular System DDocument19 pagesCardiovascular System DVarun B RenukappaPas encore d'évaluation
- Chapter 7 Cellular RespirationDocument47 pagesChapter 7 Cellular Respirationwickedbiology101Pas encore d'évaluation
- Introduction To The Body As A WholeDocument29 pagesIntroduction To The Body As A Wholekhizer hayatPas encore d'évaluation
- The Biological Perspective 1Document18 pagesThe Biological Perspective 1Radhey SurvePas encore d'évaluation
- Module 5 Pregancy - Maternal Changes 2023Document14 pagesModule 5 Pregancy - Maternal Changes 2023Berlon LacsonPas encore d'évaluation
- Ebf 303 Environmental and Animal PhysiologyDocument61 pagesEbf 303 Environmental and Animal PhysiologyAngela BrownPas encore d'évaluation
- Amino AcidsDocument18 pagesAmino AcidsShreesh MohanPas encore d'évaluation
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessD'Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessÉvaluation : 4 sur 5 étoiles4/5 (33)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseD'EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseÉvaluation : 4.5 sur 5 étoiles4.5/5 (52)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsD'EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisD'EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityD'EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityÉvaluation : 4.5 sur 5 étoiles4.5/5 (6)
- Who's in Charge?: Free Will and the Science of the BrainD'EverandWho's in Charge?: Free Will and the Science of the BrainÉvaluation : 4 sur 5 étoiles4/5 (65)
- Gut: the new and revised Sunday Times bestsellerD'EverandGut: the new and revised Sunday Times bestsellerÉvaluation : 4 sur 5 étoiles4/5 (393)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeD'EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifePas encore d'évaluation
- Tales from Both Sides of the Brain: A Life in NeuroscienceD'EverandTales from Both Sides of the Brain: A Life in NeuroscienceÉvaluation : 3 sur 5 étoiles3/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedD'EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedÉvaluation : 4 sur 5 étoiles4/5 (11)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldD'EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldÉvaluation : 4 sur 5 étoiles4/5 (597)
- Seven and a Half Lessons About the BrainD'EverandSeven and a Half Lessons About the BrainÉvaluation : 4 sur 5 étoiles4/5 (111)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouD'EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouÉvaluation : 4.5 sur 5 étoiles4.5/5 (62)
- Good Without God: What a Billion Nonreligious People Do BelieveD'EverandGood Without God: What a Billion Nonreligious People Do BelieveÉvaluation : 4 sur 5 étoiles4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorD'EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorPas encore d'évaluation
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionD'EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionÉvaluation : 4 sur 5 étoiles4/5 (812)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessD'EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessÉvaluation : 4 sur 5 étoiles4/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesD'EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesÉvaluation : 4.5 sur 5 étoiles4.5/5 (397)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceD'EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceÉvaluation : 4.5 sur 5 étoiles4.5/5 (517)
- Remnants of Ancient Life: The New Science of Old FossilsD'EverandRemnants of Ancient Life: The New Science of Old FossilsÉvaluation : 3 sur 5 étoiles3/5 (3)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignD'EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignÉvaluation : 4 sur 5 étoiles4/5 (19)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemD'EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemÉvaluation : 4.5 sur 5 étoiles4.5/5 (116)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomD'EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomÉvaluation : 4 sur 5 étoiles4/5 (216)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindD'EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindÉvaluation : 4.5 sur 5 étoiles4.5/5 (93)
- Civilized To Death: The Price of ProgressD'EverandCivilized To Death: The Price of ProgressÉvaluation : 4.5 sur 5 étoiles4.5/5 (215)