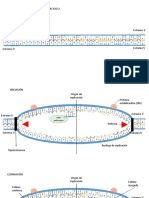
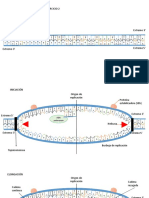
Vous aimerez peut-être aussi
- Clase 12 RsptasDocument2 pagesClase 12 RsptasalfredoPas encore d'évaluation
- Adn ReDocument4 pagesAdn RealfredoPas encore d'évaluation
- Adn ReDocument4 pagesAdn RealfredoPas encore d'évaluation
- UNIDAD 5 Autonomía y Responsabilidad IndividualDocument3 pagesUNIDAD 5 Autonomía y Responsabilidad IndividualJocelyn Aranda RabanalPas encore d'évaluation
- Clase 12Document40 pagesClase 12alfredoPas encore d'évaluation
- Helicobacter Pylori: y Enfermedades RelacionadasDocument5 pagesHelicobacter Pylori: y Enfermedades RelacionadasalfredoPas encore d'évaluation
- Clase 12 RsptasDocument2 pagesClase 12 RsptasalfredoPas encore d'évaluation
- 1ro - Plan Lector - 1ra Actividad de DiciembreDocument1 page1ro - Plan Lector - 1ra Actividad de DiciembrealfredoPas encore d'évaluation
- EjerciciosDocument2 pagesEjerciciosalfredoPas encore d'évaluation
- Clase 9. Naturaleza Del Gen y El Genoma 2020-2Document24 pagesClase 9. Naturaleza Del Gen y El Genoma 2020-2alfredoPas encore d'évaluation
- 1ro - Plan Lector - 1ra Actividad de DiciembreDocument1 page1ro - Plan Lector - 1ra Actividad de DiciembrealfredoPas encore d'évaluation
- Adn ReDocument4 pagesAdn RealfredoPas encore d'évaluation
- Polimorfismo de MHC-IIDocument1 pagePolimorfismo de MHC-IIalfredoPas encore d'évaluation
- HHDocument8 pagesHHalfredoPas encore d'évaluation
- MurrayDocument1 pageMurrayalfredoPas encore d'évaluation
- FiorellaDocument7 pagesFiorellaalfredoPas encore d'évaluation
- Guerra ADocument2 pagesGuerra AalfredoPas encore d'évaluation
- 1 PDFDocument10 pages1 PDFalfredoPas encore d'évaluation
- La Sexualidad AnaliticaDocument1 pageLa Sexualidad AnaliticaalfredoPas encore d'évaluation
- Embarazo incipiente 5 semanas sin latido cardiacoDocument3 pagesEmbarazo incipiente 5 semanas sin latido cardiacoalfredoPas encore d'évaluation
- Plan de Trabajo de FilosofiaDocument5 pagesPlan de Trabajo de FilosofiaalfredoPas encore d'évaluation
- Analisis CompetititvoDocument2 pagesAnalisis CompetititvoalfredoPas encore d'évaluation
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoPas encore d'évaluation
- Contaminación Del Parque AutomotorDocument29 pagesContaminación Del Parque Automotorwillgeiner100% (5)
- Ensayo FinalDocument11 pagesEnsayo FinalalfredoPas encore d'évaluation
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoPas encore d'évaluation
- Contaminación Del Parque AutomotorDocument29 pagesContaminación Del Parque Automotorwillgeiner100% (5)
- Articulo de OpinionDocument5 pagesArticulo de OpinionalfredoPas encore d'évaluation
- Informe EstadisticoDocument30 pagesInforme EstadisticoalfredoPas encore d'évaluation
- Informe Estadistico TerminadoDocument31 pagesInforme Estadistico TerminadoalfredoPas encore d'évaluation
- Hepatitis Viral AgudaDocument56 pagesHepatitis Viral AgudaWillyam MaqueraPas encore d'évaluation
- Tratamiento de Las Enfermedades Hepáticas y BiliaresDocument506 pagesTratamiento de Las Enfermedades Hepáticas y BiliaresDavid MolinaPas encore d'évaluation
- Hepatitis ViralesDocument14 pagesHepatitis ViralesGlenda VizcaínoPas encore d'évaluation
- Mad Rid - Nº71Document65 pagesMad Rid - Nº71José Ramón SndVgPas encore d'évaluation
- Pre Internado 2021 - Villamemo PediatríaDocument23 pagesPre Internado 2021 - Villamemo PediatríaANTOLINO GENARO ROSALES PIJOPas encore d'évaluation
- AntiviralesDocument15 pagesAntiviralesCrista Villatoro100% (1)
- 23 - Virus Hepatotropos PDFDocument9 pages23 - Virus Hepatotropos PDFPaco PérePas encore d'évaluation
- Pabellon o Bandera Nacional de Guatemala. - Tipos de Banderas Que Han Existido A Lo Largo de La Historia. - Virus, Bacterias y EjemplosDocument9 pagesPabellon o Bandera Nacional de Guatemala. - Tipos de Banderas Que Han Existido A Lo Largo de La Historia. - Virus, Bacterias y EjemplosSelvyn González100% (1)
- Resumen RAE 4 IctericiaDocument4 pagesResumen RAE 4 IctericiaJuan Diego Barragan PinillaPas encore d'évaluation
- Hepatitis BDocument15 pagesHepatitis BIrving PlazaPas encore d'évaluation
- Universidad Autonoma "Juan Misael Saracho"Document70 pagesUniversidad Autonoma "Juan Misael Saracho"LuisAlbertoChipanaMamani100% (1)
- Hepatitis BDocument2 pagesHepatitis BJesus Espiritu Morales SuyonPas encore d'évaluation
- Virus de la hepatitis: causas, síntomas y prevenciónDocument30 pagesVirus de la hepatitis: causas, síntomas y prevenciónValeria Córdova ReyesPas encore d'évaluation
- Hepatitis BDocument13 pagesHepatitis BOswaldo MonsalvePas encore d'évaluation
- Hepatitis AbcdeDocument20 pagesHepatitis Abcdesaulcarvajal23Pas encore d'évaluation
- Evaluación paciente trasplante renalDocument18 pagesEvaluación paciente trasplante renalIgnacio VarelaPas encore d'évaluation
- Arancel OchagavíaDocument13 pagesArancel OchagavíaJosé López RodríguezPas encore d'évaluation
- Plan de Tesis (Avance) MedicinaDocument43 pagesPlan de Tesis (Avance) Medicinaangel moe 777Pas encore d'évaluation
- Hepatitis ViralesDocument20 pagesHepatitis ViralesAdela MejíaPas encore d'évaluation
- RM Norma Tecnica Hepatitis BDocument24 pagesRM Norma Tecnica Hepatitis BArmandina PerezPas encore d'évaluation
- Conversion ResultDocument70 pagesConversion ResultFrancisco Javier Muñoz FonttzPas encore d'évaluation
- Virus de Las HepatitisDocument86 pagesVirus de Las HepatitisALEXANDRA INAGANPas encore d'évaluation
- Virus HepatotroposDocument94 pagesVirus HepatotroposJohanna MarchanPas encore d'évaluation
- Capitulo Hepatitis-NegroniDocument12 pagesCapitulo Hepatitis-NegroniAna valeria MuñozPas encore d'évaluation
- Padron Its 2019 JulioDocument67 pagesPadron Its 2019 JulioWilfredo Mateo YantasPas encore d'évaluation
- Hepatitis BDocument8 pagesHepatitis BErika Arlete Gaspar ValenciaPas encore d'évaluation
- Virus oncogénicos y cáncer humanoDocument5 pagesVirus oncogénicos y cáncer humanoGualterney AmarilioPas encore d'évaluation
- Virología Hepatitis BDocument13 pagesVirología Hepatitis BEunice AriasPas encore d'évaluation
- Higado Patologia de Robbins y CotranDocument12 pagesHigado Patologia de Robbins y CotranKarla Ariza Hernandez60% (5)
- Hepatitis Viral: Agentes, Transmisión y Cuadros ClínicosDocument22 pagesHepatitis Viral: Agentes, Transmisión y Cuadros ClínicosScarlett Sepulveda100% (1)