Vous aimerez peut-être aussi
- Environmental and Health Impacts of Air PollutionDocument13 pagesEnvironmental and Health Impacts of Air PollutionJUAN SEBASTIAN ARISTIZABAL APOLINARPas encore d'évaluation
- Pollution The SDocument15 pagesPollution The SWasil SaipodenPas encore d'évaluation
- Meliana 12345Document11 pagesMeliana 12345KikiaraPas encore d'évaluation
- Air Pollution Effects On The Gut MicrobiotaDocument6 pagesAir Pollution Effects On The Gut MicrobiotaTrydawPas encore d'évaluation
- Jurnal Pembanding (Internasional)Document8 pagesJurnal Pembanding (Internasional)Elisabeth Anggelika NahampunPas encore d'évaluation
- EHP1699.altDocument11 pagesEHP1699.altkunkkonkPas encore d'évaluation
- Environmental Research: Review ArticleDocument7 pagesEnvironmental Research: Review ArticleMaria Jose Altamirano CastroPas encore d'évaluation
- Popovic - Et.al.2019. A Systematic Review and Critical Aprasial of Epidemiological Studies On Outdoor Air Polloution and TB OutcomesDocument13 pagesPopovic - Et.al.2019. A Systematic Review and Critical Aprasial of Epidemiological Studies On Outdoor Air Polloution and TB OutcomesmichaellouisgrecoPas encore d'évaluation
- EScholarship UC Item 0fq156mxDocument11 pagesEScholarship UC Item 0fq156mxDian MaulidaPas encore d'évaluation
- Prata 2019Document32 pagesPrata 2019shefali bageriaPas encore d'évaluation
- Air PollutionDocument30 pagesAir Pollutionrudra royPas encore d'évaluation
- 11 - Chapter 1Document14 pages11 - Chapter 1Mohammad Zamir TaqwaPas encore d'évaluation
- A Review by The Forum of International Respiratory SocietiesEnvironmental Committee, Part 1Document8 pagesA Review by The Forum of International Respiratory SocietiesEnvironmental Committee, Part 1lidysPas encore d'évaluation
- Environmental Research: Jimyung Park, Hyung-Jun Kim, Chang-Hoon Lee, Chang Hyun Lee, Hyun Woo LeeDocument8 pagesEnvironmental Research: Jimyung Park, Hyung-Jun Kim, Chang-Hoon Lee, Chang Hyun Lee, Hyun Woo LeeJoaquim Lorenzetti BrancoPas encore d'évaluation
- OECD Covid Env Health Brief April 2020Document13 pagesOECD Covid Env Health Brief April 2020Nidia PontePas encore d'évaluation
- Respirology - 2012 - SIERRA VARGAS - Air Pollution Impact and PreventionDocument8 pagesRespirology - 2012 - SIERRA VARGAS - Air Pollution Impact and PreventionSorina ElenaPas encore d'évaluation
- Fpubh 11 1140786Document18 pagesFpubh 11 1140786Gianina CraiaPas encore d'évaluation
- Seasonal Associations Between Air Pollutant - 2023 - International Journal of HyDocument8 pagesSeasonal Associations Between Air Pollutant - 2023 - International Journal of HyDana MateiPas encore d'évaluation
- Potential Roles of Air Pollutants On The Induction and Aggr - 2023 - EnvironmentDocument14 pagesPotential Roles of Air Pollutants On The Induction and Aggr - 2023 - EnvironmentDana MateiPas encore d'évaluation
- Tuberculos MedicamentoDocument9 pagesTuberculos MedicamentoTatiana Amaya GomezPas encore d'évaluation
- Biomedical Journal: SciencedirectDocument22 pagesBiomedical Journal: SciencedirectmsdhaniaPas encore d'évaluation
- Fine Particle Environmental Pollution and Cardiovascular DiseasesDocument5 pagesFine Particle Environmental Pollution and Cardiovascular Diseasespao.pearanda1998Pas encore d'évaluation
- Liu 2018Document11 pagesLiu 2018Jimmy NarvaezPas encore d'évaluation
- Air Pollution Project ProposalDocument4 pagesAir Pollution Project ProposalQuenlay KP Wangchukk50% (2)
- Air Pollution and Lung HealthDocument8 pagesAir Pollution and Lung HealthSherifaPas encore d'évaluation
- Interactions of Climate Change, Air Pollution, and Human HealthDocument8 pagesInteractions of Climate Change, Air Pollution, and Human Healthchandgagan60Pas encore d'évaluation
- 10 1002@jat 3940Document15 pages10 1002@jat 3940Nadya AlizaPas encore d'évaluation
- Fpubh 10 882569Document13 pagesFpubh 10 882569Katalin BodorPas encore d'évaluation
- PS Effect in Pulmonary Cells HumanDocument40 pagesPS Effect in Pulmonary Cells Humanmegha deyPas encore d'évaluation
- Building Resilient Cities Climate Change and Health Interlinkages in The Planning of Public SpacesDocument22 pagesBuilding Resilient Cities Climate Change and Health Interlinkages in The Planning of Public Spaceslailanurhikmah07Pas encore d'évaluation
- Baboolal Et Al 2019 API TrinidadDocument16 pagesBaboolal Et Al 2019 API TrinidadHema BaboolalPas encore d'évaluation
- Human Health PDFDocument29 pagesHuman Health PDFJay Mark RepontePas encore d'évaluation
- PM2 5Document23 pagesPM2 5Trọng KhảiPas encore d'évaluation
- Climate Change PaperDocument13 pagesClimate Change Paperapi-531666073Pas encore d'évaluation
- Content ServerDocument7 pagesContent ServerYENIFER SANCHEZ NAVARROPas encore d'évaluation
- Analysis of Human Health Impacts On Air Pollution at Global LevelDocument7 pagesAnalysis of Human Health Impacts On Air Pollution at Global LevelSanjoy MajiPas encore d'évaluation
- The Effect of Air Pollution in The Environment and The Public HealthDocument6 pagesThe Effect of Air Pollution in The Environment and The Public Healthlyne100% (1)
- INTRODUCTIONDocument14 pagesINTRODUCTIONmoses samuelPas encore d'évaluation
- Ijerph 19 07511Document22 pagesIjerph 19 07511Katalin BodorPas encore d'évaluation
- Nepal Journal of Biotechnology - Vol. 9 No. 1 (2021)Document12 pagesNepal Journal of Biotechnology - Vol. 9 No. 1 (2021)mpd26Pas encore d'évaluation
- Ijerph 15 00780 PDFDocument44 pagesIjerph 15 00780 PDFSanyamJainPas encore d'évaluation
- Cata Mauria Case Study EcoDocument6 pagesCata Mauria Case Study EcoLalaine RideraPas encore d'évaluation
- Global Mortality From Outdoor Fine Particle by FossilDocument8 pagesGlobal Mortality From Outdoor Fine Particle by FossilAHMAD BALFAQIHPas encore d'évaluation
- Air Pollution Exposure and Blood Pressure: An Updated Review of The LiteratureDocument24 pagesAir Pollution Exposure and Blood Pressure: An Updated Review of The LiteratureJackson LPas encore d'évaluation
- Research Article Exposure To Air Pollution Exacerbates Inflammation in Rats With Preexisting COPDDocument12 pagesResearch Article Exposure To Air Pollution Exacerbates Inflammation in Rats With Preexisting COPDĐạt NguyễnPas encore d'évaluation
- Assessment of Airborne Bacteria and Fungi in Various University Indoor Environment in Chang'an University ChinaDocument11 pagesAssessment of Airborne Bacteria and Fungi in Various University Indoor Environment in Chang'an University ChinaGratia TomasoaPas encore d'évaluation
- Using Regression Models To Predict Death Caused by Ambient Ozone Pollution (AOP) in The United StatesDocument18 pagesUsing Regression Models To Predict Death Caused by Ambient Ozone Pollution (AOP) in The United StatesInternational Journal of Innovative Science and Research Technology100% (1)
- Effects of Particulate Matter On Allergic Respiratory Diseases.Document8 pagesEffects of Particulate Matter On Allergic Respiratory Diseases.wawa chenPas encore d'évaluation
- Environment-Pathogen Interaction in Plant Diseases: Review ArticleDocument8 pagesEnvironment-Pathogen Interaction in Plant Diseases: Review ArticleRajat SharmaPas encore d'évaluation
- Patel Et Al 2020 - Poluentes Emergentes Amb AquaticoDocument15 pagesPatel Et Al 2020 - Poluentes Emergentes Amb AquaticogddezorziPas encore d'évaluation
- 2021 - Effects of PM10 Andweather On Respiratory and CardiovascularDocument21 pages2021 - Effects of PM10 Andweather On Respiratory and CardiovascularBodor ZsoltPas encore d'évaluation
- 5) Cohen2017Document12 pages5) Cohen2017Chintrai ThavonlunPas encore d'évaluation
- Pollution and Respiratory Disease: Can Diet or Supplements Help? A ReviewDocument14 pagesPollution and Respiratory Disease: Can Diet or Supplements Help? A ReviewEgy SunandaPas encore d'évaluation
- Anu Chapter 1Document7 pagesAnu Chapter 1Arun AGPas encore d'évaluation
- 1 s2.0 S0147651320307491 MainDocument12 pages1 s2.0 S0147651320307491 MainmuhammedPas encore d'évaluation
- Seasonal Variation in Culturable Bioaerosols in A Wastewater Treatment PlantDocument14 pagesSeasonal Variation in Culturable Bioaerosols in A Wastewater Treatment Plantcharvi shinyPas encore d'évaluation
- Pollution in UKDocument13 pagesPollution in UKHassan TariqPas encore d'évaluation
- Risk Assessment of Exposure To Volatile Organic Compounds in Different Indoor EnvironmentsDocument10 pagesRisk Assessment of Exposure To Volatile Organic Compounds in Different Indoor EnvironmentsiprateekPas encore d'évaluation
- Health Apprehensive Level of Canteen Workers in Western Mindanao State University On Carbon MonoxideDocument10 pagesHealth Apprehensive Level of Canteen Workers in Western Mindanao State University On Carbon Monoxideruth lopezPas encore d'évaluation
- Inhibition Killing NPDocument8 pagesInhibition Killing NPSuzan MatarPas encore d'évaluation
- 2020 Evalution FormDocument1 page2020 Evalution FormSuzan MatarPas encore d'évaluation
- Nehaya s41467-019-08719-8Document12 pagesNehaya s41467-019-08719-8Suzan MatarPas encore d'évaluation
- Polyclnal Antibody ProductionDocument24 pagesPolyclnal Antibody ProductionSuzan MatarPas encore d'évaluation
- Biomaterials: Kyle G. Battiston, Jane W.C. Cheung, Devika Jain, J. Paul SanterreDocument12 pagesBiomaterials: Kyle G. Battiston, Jane W.C. Cheung, Devika Jain, J. Paul SanterreSuzan MatarPas encore d'évaluation
- Basic Problems of Serological Laboratory DiagnosisDocument37 pagesBasic Problems of Serological Laboratory DiagnosisSuzan MatarPas encore d'évaluation
- WHO Lab Quality Management SystemDocument246 pagesWHO Lab Quality Management SystemMigori Art100% (1)
- Medical Terminology Learning Outcome 1: Apply The Rules For Construction and Analysis of Medical TermsDocument15 pagesMedical Terminology Learning Outcome 1: Apply The Rules For Construction and Analysis of Medical TermsSuzan MatarPas encore d'évaluation
- M STC Medical Terminology Web InfoDocument3 pagesM STC Medical Terminology Web InfoSuzan MatarPas encore d'évaluation
- Normal Flora of The Human BodyDocument4 pagesNormal Flora of The Human BodySuzan MatarPas encore d'évaluation
- Integrated Management of Childhood Illness: Child Aged 2 Months Up To 5 YearsDocument44 pagesIntegrated Management of Childhood Illness: Child Aged 2 Months Up To 5 YearsJulio Iñigo López100% (22)
- Community Acquired Pneumonia 2Document22 pagesCommunity Acquired Pneumonia 2carosegurallPas encore d'évaluation
- CD Respi 30 ADocument3 pagesCD Respi 30 ANae OrdanozoPas encore d'évaluation
- NCP PneumoniaDocument6 pagesNCP PneumoniaMerliah LofrancoPas encore d'évaluation
- 500 CRT Practice QuestionsDocument138 pages500 CRT Practice QuestionsCPD MAS50% (2)
- Physical Education 12 Pneumonia: To Be Reported By: Alawi, Lhamiah M. Casano, Diana Elizabeth C. Kinoshita, Ryou GDocument21 pagesPhysical Education 12 Pneumonia: To Be Reported By: Alawi, Lhamiah M. Casano, Diana Elizabeth C. Kinoshita, Ryou GRyou KinoshitaPas encore d'évaluation
- Acute Respiratory InfectionDocument9 pagesAcute Respiratory InfectionStephen FoxPas encore d'évaluation
- Guia 2023 de Manejo de SdraDocument9 pagesGuia 2023 de Manejo de SdraBrian Antonio Veramatos LopezPas encore d'évaluation
- High Resolution Computed Tomography ManifestationsDocument6 pagesHigh Resolution Computed Tomography ManifestationseugeniaPas encore d'évaluation
- Killer Whales (Orcas) That Died in Captivity by NameDocument6 pagesKiller Whales (Orcas) That Died in Captivity by NameThe Orca Project Corp100% (8)
- Pediatric Community-Acquired Pneumonia: CC Trisha Pamela OquendoDocument70 pagesPediatric Community-Acquired Pneumonia: CC Trisha Pamela OquendoJessa MaePas encore d'évaluation
- CAFL DatabaseDocument178 pagesCAFL Databasetonym35100% (1)
- Aspiration PneumoniaDocument22 pagesAspiration PneumoniaAya AlamsjahPas encore d'évaluation
- IMNCI Students Handbook PDFDocument93 pagesIMNCI Students Handbook PDFAamira Qureshi100% (4)
- Polyclinic AllDocument92 pagesPolyclinic AllFatima ShahPas encore d'évaluation
- Summary Test 2. Diseases of Respiratory Tract in ChildrenDocument14 pagesSummary Test 2. Diseases of Respiratory Tract in Childrenprashant singh0% (1)
- Respiratory DisordersDocument48 pagesRespiratory DisordersCalimlim KimPas encore d'évaluation
- Pneumonia: Ma. Concepcion Badar Liezl Joy Stanfa Marilyn BuctuanDocument29 pagesPneumonia: Ma. Concepcion Badar Liezl Joy Stanfa Marilyn Buctuanarl777Pas encore d'évaluation
- M6 Bacteriology Pre Task - CaparasDocument1 pageM6 Bacteriology Pre Task - CaparasGretta CaparasPas encore d'évaluation
- 5G Radiation and The COVID-19 Pandemic: Coincidence or Causal RelationshipDocument3 pages5G Radiation and The COVID-19 Pandemic: Coincidence or Causal RelationshipMartin L. Pall100% (10)
- Hiv-Sle Case StudyDocument8 pagesHiv-Sle Case StudycathyPas encore d'évaluation
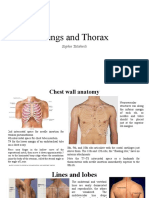
- Lungs and Thorax: Sophie TatishviliDocument31 pagesLungs and Thorax: Sophie TatishviliAstha Tusharbhai PatelPas encore d'évaluation
- 3 Infeksi Saluran Pernafasan Akut (ISPA) PediatricsDocument35 pages3 Infeksi Saluran Pernafasan Akut (ISPA) PediatricsCharisma Tiara RessyaPas encore d'évaluation
- Transient Tachypnea of The NewbornDocument14 pagesTransient Tachypnea of The NewbornMarielaTessyGonzalesParedesPas encore d'évaluation
- Cluster 5Document12 pagesCluster 5st. hyacinth radioPas encore d'évaluation
- Mock 2Document40 pagesMock 2Syali Sasidharan100% (2)
- Confidential GlaxoSmithKline Document Shows 36 Infant Deaths Following VaccinationDocument1 271 pagesConfidential GlaxoSmithKline Document Shows 36 Infant Deaths Following Vaccinationdiligentpurpose100% (2)
- Respiratory Tract Infection Pharmacotherapy RevisedDocument82 pagesRespiratory Tract Infection Pharmacotherapy Revisedchuye alemayehuPas encore d'évaluation
- q1121 PDFDocument8 pagesq1121 PDFKijendran Chandra SekarPas encore d'évaluation
- Respiratory SystemDocument9 pagesRespiratory Systemtheglobalnursing89% (9)
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseD'EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseÉvaluation : 3.5 sur 5 étoiles3.5/5 (69)
- The Other End of the Leash: Why We Do What We Do Around DogsD'EverandThe Other End of the Leash: Why We Do What We Do Around DogsÉvaluation : 5 sur 5 étoiles5/5 (65)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildD'EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildÉvaluation : 4.5 sur 5 étoiles4.5/5 (44)
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessD'EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessPas encore d'évaluation
- Spoiled Rotten America: Outrages of Everyday LifeD'EverandSpoiled Rotten America: Outrages of Everyday LifeÉvaluation : 3 sur 5 étoiles3/5 (19)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionD'EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionÉvaluation : 4 sur 5 étoiles4/5 (812)
- The Best American Science And Nature Writing 2021D'EverandThe Best American Science And Nature Writing 2021Évaluation : 4 sur 5 étoiles4/5 (8)
- The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorD'EverandThe Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorÉvaluation : 4.5 sur 5 étoiles4.5/5 (137)
- When You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsD'EverandWhen You Find Out the World Is Against You: And Other Funny Memories About Awful MomentsÉvaluation : 3.5 sur 5 étoiles3.5/5 (13)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldD'EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldÉvaluation : 4 sur 5 étoiles4/5 (597)
- Fire Season: Field Notes from a Wilderness LookoutD'EverandFire Season: Field Notes from a Wilderness LookoutÉvaluation : 4 sur 5 étoiles4/5 (142)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingD'EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingÉvaluation : 5 sur 5 étoiles5/5 (5)
- World of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsD'EverandWorld of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsÉvaluation : 4 sur 5 étoiles4/5 (223)
- Come Back, Como: Winning the Heart of a Reluctant DogD'EverandCome Back, Como: Winning the Heart of a Reluctant DogÉvaluation : 3.5 sur 5 étoiles3.5/5 (10)
- Remnants of Ancient Life: The New Science of Old FossilsD'EverandRemnants of Ancient Life: The New Science of Old FossilsÉvaluation : 3 sur 5 étoiles3/5 (3)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignD'EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignÉvaluation : 4 sur 5 étoiles4/5 (19)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldD'EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldÉvaluation : 4.5 sur 5 étoiles4.5/5 (18)
- The Mind of Plants: Narratives of Vegetal IntelligenceD'EverandThe Mind of Plants: Narratives of Vegetal IntelligenceÉvaluation : 4.5 sur 5 étoiles4.5/5 (12)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingD'EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (35)
- The Hawk's Way: Encounters with Fierce BeautyD'EverandThe Hawk's Way: Encounters with Fierce BeautyÉvaluation : 4.5 sur 5 étoiles4.5/5 (19)