Vous aimerez peut-être aussi
- Regulacion Genica en ProcariotasDocument10 pagesRegulacion Genica en ProcariotasMaría Juliana Rojas MoncadaPas encore d'évaluation
- Genes y Control de La Expresión GenéticaDocument12 pagesGenes y Control de La Expresión GenéticaYAHIZIRI JANAIL VENTURA SAMAYOAPas encore d'évaluation
- Expresión GénicaDocument48 pagesExpresión GénicaAshley BrigittePas encore d'évaluation
- Regulacion Genica ResumenDocument3 pagesRegulacion Genica ResumenEstrella MartínezPas encore d'évaluation
- Operon LactosaDocument19 pagesOperon LactosaJohn JuradoPas encore d'évaluation
- Regulación de La Expresión GeneticaDocument61 pagesRegulación de La Expresión Geneticalmgarcia204Pas encore d'évaluation
- Informe 4 Genetica Parte IsaDocument4 pagesInforme 4 Genetica Parte IsaIsaias Lopez EstradaPas encore d'évaluation
- Castro Ocampo Viridiana. Regulación Genética de Procariontes.Document44 pagesCastro Ocampo Viridiana. Regulación Genética de Procariontes.Viridiana Castro OcampoPas encore d'évaluation
- Ensayo Biologia Del Desarrollo AnimalDocument19 pagesEnsayo Biologia Del Desarrollo AnimalNADIAPas encore d'évaluation
- 12 Control de La Expresion Genica Lac TRPDocument30 pages12 Control de La Expresion Genica Lac TRPWilliam Nicolás Bacca NovoaPas encore d'évaluation
- Operon LactosaDocument52 pagesOperon LactosaAmbar GabrielaPas encore d'évaluation
- OPERONDocument15 pagesOPERONGineth MaderaPas encore d'évaluation
- Regulación de La Expresión GenéticaDocument6 pagesRegulación de La Expresión GenéticaJorge Carlos Vazquez SanchezPas encore d'évaluation
- Regulacion Genica en Procariotas PDFDocument27 pagesRegulacion Genica en Procariotas PDFMarleni Rivera CastromontePas encore d'évaluation
- Control de La Función Génica y Actividad Bioquímica ResumenDocument3 pagesControl de La Función Génica y Actividad Bioquímica ResumenJairoCarreraPas encore d'évaluation
- Regulación de La Expresión Génica en Bacterias!!Document27 pagesRegulación de La Expresión Génica en Bacterias!!Laura Téllez LópezPas encore d'évaluation
- Tema 8-Bloque I-Regulacion GeneticaDocument8 pagesTema 8-Bloque I-Regulacion GeneticaUriel Rosas HuertaPas encore d'évaluation
- Regulacion de La Expresion Genetica en BacteriaDocument27 pagesRegulacion de La Expresion Genetica en BacteriaMaggie Tapia ZúñigaPas encore d'évaluation
- Regulación en EucariotasDocument13 pagesRegulación en EucariotasDaryl TinjacaPas encore d'évaluation
- OperonesDocument31 pagesOperonesDiego A Brito DominguezPas encore d'évaluation
- Ensayo Operones y ArniDocument9 pagesEnsayo Operones y ArniAntonio Tenorio GonzalezPas encore d'évaluation
- Trabajo. Operon Lactosa.Document9 pagesTrabajo. Operon Lactosa.Jhonier Jesús Ruiz PèrezPas encore d'évaluation
- Experimento de Beadle y TatumDocument7 pagesExperimento de Beadle y TatumJuli SaplanaPas encore d'évaluation
- Operon GuiaDocument16 pagesOperon GuiaEugaldePas encore d'évaluation
- Sistema de Expresión de Genes en ProcariotasDocument2 pagesSistema de Expresión de Genes en ProcariotasValeria GonzálezPas encore d'évaluation
- Regulacion Procariotas 2024Document64 pagesRegulacion Procariotas 2024Brayan MejíasPas encore d'évaluation
- Regulación GénicaDocument40 pagesRegulación GénicaISBELIAC7Pas encore d'évaluation
- Elementos Del OperónDocument16 pagesElementos Del OperónKristian CobaPas encore d'évaluation
- Regulación A Nivel de DnaDocument5 pagesRegulación A Nivel de DnaMaría AhumadaPas encore d'évaluation
- Taller 3 Tecnicas de Biologia MolecularDocument5 pagesTaller 3 Tecnicas de Biologia MolecularJulieta PedrouzoPas encore d'évaluation
- 3.3. Regulación A Nivel ADNDocument12 pages3.3. Regulación A Nivel ADNJesusEnrikeSerranoPas encore d'évaluation
- Exposición Biología MolecularDocument5 pagesExposición Biología MolecularDIANA TALAVERA ZERMEÑOPas encore d'évaluation
- Nucleo en Una Celula EucariotaDocument3 pagesNucleo en Una Celula EucariotaIveth MelendrezPas encore d'évaluation
- REGULACIONDocument12 pagesREGULACIONMartinez avila Jose GaelPas encore d'évaluation
- 6) Regulacion - Genica - Con AudioDocument29 pages6) Regulacion - Genica - Con AudioNasrry Kaleb PoncePas encore d'évaluation
- Tema 7 - Regulacion de La Expresion GénicaDocument19 pagesTema 7 - Regulacion de La Expresion GénicaVicky CruzPas encore d'évaluation
- Teoria Del OperonDocument8 pagesTeoria Del OperonValeria Rz100% (1)
- Regulación de La Expresión GénicaDocument12 pagesRegulación de La Expresión GénicaJessicaSilveroPas encore d'évaluation
- RegexprgenicaDocument40 pagesRegexprgenicaMelanie Valeria Pulido SanchezPas encore d'évaluation
- 2.5 OperonesDocument26 pages2.5 OperonesAlonso ZavalaPas encore d'évaluation
- Tema 64Document8 pagesTema 64Virginia Romero CarrascoPas encore d'évaluation
- OperonDocument32 pagesOperonfrancisco lozano gomezPas encore d'évaluation
- UD-Genética Molecular II MODIFICADODocument8 pagesUD-Genética Molecular II MODIFICADODavidPas encore d'évaluation
- Regulacion Genica en ProcariotasDocument27 pagesRegulacion Genica en ProcariotasMarleni Rivera CastromontePas encore d'évaluation
- Semana 13 - TeoriaDocument20 pagesSemana 13 - TeoriajoselinPas encore d'évaluation
- Control de La Expresión GénicaDocument24 pagesControl de La Expresión GénicaCamila ObregonPas encore d'évaluation
- MaterialesDocument13 pagesMaterialesYANETHPas encore d'évaluation
- Taller de Genetica MicrobianaDocument8 pagesTaller de Genetica Microbianacarlos vargasPas encore d'évaluation
- Control Genético Del DesarrolloDocument8 pagesControl Genético Del DesarrolloLaury CustodioPas encore d'évaluation
- GENETICADocument14 pagesGENETICAYANETHPas encore d'évaluation
- Regulación TranscripcionalDocument31 pagesRegulación TranscripcionalFermín Armando Ledezma VelázquezPas encore d'évaluation
- Regulación GénicaDocument3 pagesRegulación GénicaFrancisco Javier F. LeónPas encore d'évaluation
- Genetica BacterianaDocument103 pagesGenetica BacterianaMaryGadeaPas encore d'évaluation
- 4°BIO PD1 Guía SD0SDA3TAREA12 1 Al 5 06Document4 pages4°BIO PD1 Guía SD0SDA3TAREA12 1 Al 5 06Francisca HermosillaPas encore d'évaluation
- OperonDocument6 pagesOperonLuz AndreaPas encore d'évaluation
- Relacion Existente Entre Los Genes y La Diferenciacion Celular Que Ocurre Durante El Desarrollo EmbrionarioDocument10 pagesRelacion Existente Entre Los Genes y La Diferenciacion Celular Que Ocurre Durante El Desarrollo EmbrionarioCristinaGonzalezPas encore d'évaluation
- Operon LacDocument6 pagesOperon LacJerlys Quijano.Pas encore d'évaluation
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónD'EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónPas encore d'évaluation
- Epigenética para intermedios. La exploración más completa del impacto práctico, social y ético del ADN en nuestra sociedad y nuestro mundoD'EverandEpigenética para intermedios. La exploración más completa del impacto práctico, social y ético del ADN en nuestra sociedad y nuestro mundoPas encore d'évaluation
- James DouglasDocument3 pagesJames DouglasBenjamin Padilla ParodiPas encore d'évaluation
- CachicheDocument2 pagesCachicheBenjamin Padilla ParodiPas encore d'évaluation
- AneurismaDocument5 pagesAneurismaBenjamin Padilla ParodiPas encore d'évaluation
- Scuderia FerrariDocument2 pagesScuderia FerrariBenjamin Padilla ParodiPas encore d'évaluation
- Resumen EsterilizaciónDocument8 pagesResumen EsterilizaciónBenjamin Padilla ParodiPas encore d'évaluation
- Pruebas Usadas para Diagnosticar y Clasificar La Leucemia WordDocument19 pagesPruebas Usadas para Diagnosticar y Clasificar La Leucemia WordBenjamin Padilla ParodiPas encore d'évaluation
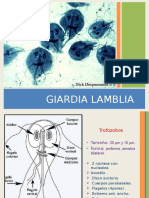
- GiardiaDocument7 pagesGiardiaBenjamin Padilla ParodiPas encore d'évaluation
- Especial EsDocument5 pagesEspecial EsBenjamin Padilla ParodiPas encore d'évaluation
- Himenolepiosis y DipilidiosisDocument24 pagesHimenolepiosis y DipilidiosisBenjamin Padilla ParodiPas encore d'évaluation
- Laboratorio 1 - Transduccion GeneralDocument16 pagesLaboratorio 1 - Transduccion Generaljorge fravega ammannPas encore d'évaluation
- Fichas FinalesDocument24 pagesFichas FinalesMadhy MartinezPas encore d'évaluation
- Matematicas y MicroscopioDocument29 pagesMatematicas y MicroscopioEduardo Lulichac RojasPas encore d'évaluation
- Reproducción en Organismos Sencillo1Document6 pagesReproducción en Organismos Sencillo1diana isabelPas encore d'évaluation
- Genetica MicrobianaDocument26 pagesGenetica MicrobianaSanti MontiPas encore d'évaluation
- Resumen Del Libro MicrocosmosDocument7 pagesResumen Del Libro Microcosmosloli100% (1)
- ClassesallDocument44 pagesClassesallEstefania AguilarPas encore d'évaluation
- EUI Microbiología 2020Document12 pagesEUI Microbiología 2020Cristhian Daniel Chachaque TiconaPas encore d'évaluation
- Apuntes Segundo Parcial Biologia MolecularDocument12 pagesApuntes Segundo Parcial Biologia MolecularDiego LaraPas encore d'évaluation
- 0apuntes MicroDocument135 pages0apuntes Microanita6208Pas encore d'évaluation
- Ejercicios Pau Bloque IV - SolucionesDocument31 pagesEjercicios Pau Bloque IV - Solucionesf97yrpddvdPas encore d'évaluation
- Bloque 5 Biologia II Parte 2Document74 pagesBloque 5 Biologia II Parte 2Irvin GutiérrezPas encore d'évaluation
- Resumen Microbiologia BacteriasDocument29 pagesResumen Microbiologia BacteriasJhormanQuirozPas encore d'évaluation
- 1 Genoma ViricoDocument10 pages1 Genoma ViricoClara Silva100% (1)
- Vectores de Clonación y ExpresiónDocument42 pagesVectores de Clonación y ExpresiónJuliana MuñozPas encore d'évaluation
- Vibrio Cholerae (1) .En - Es PDFDocument14 pagesVibrio Cholerae (1) .En - Es PDFGiss EstevesPas encore d'évaluation
- Aislamiento de BacteriofagosDocument6 pagesAislamiento de BacteriofagosGiselle Escobedo CortijoPas encore d'évaluation
- Biomagnetismo Avanzado Versión EspañolDocument45 pagesBiomagnetismo Avanzado Versión EspañolozsmypPas encore d'évaluation
- ListeriaDocument38 pagesListeriasmstudenkoPas encore d'évaluation
- Vectores LambdaDocument5 pagesVectores Lambdamartasanz34100% (3)
- Maduración y Liberación Viral.Document25 pagesMaduración y Liberación Viral.Danny Suclupe Campos100% (1)
- Genetica BacterianaDocument33 pagesGenetica Bacterianamesthudiilloo100% (1)
- Genética GeneralDocument7 pagesGenética GeneralAmilcar FermínPas encore d'évaluation
- Epidemias y Salud Publica - PDF Versión 1Document77 pagesEpidemias y Salud Publica - PDF Versión 1maria4moreno-71Pas encore d'évaluation
- Aislamiento de BacteriófagosDocument5 pagesAislamiento de BacteriófagosLuis LeónPas encore d'évaluation
- Virologia 1 UsamedicDocument111 pagesVirologia 1 UsamedicLii LiiPas encore d'évaluation
- Virus RepasoDocument35 pagesVirus RepasoSofia Isabel Espinoza CarhuayalPas encore d'évaluation
- Resumen Microbiologia GralDocument37 pagesResumen Microbiologia GralDaiana Aden100% (1)
- FagosDocument49 pagesFagosMoliriPas encore d'évaluation
- 1662 3 5350 1 10 20210716Document181 pages1662 3 5350 1 10 20210716Joffre Danilo Heredia MendozaPas encore d'évaluation