Vous aimerez peut-être aussi
- Comparative Prod of Amylase Enzyme From RhizopusDocument4 pagesComparative Prod of Amylase Enzyme From RhizopusIsworo RukmiPas encore d'évaluation
- Fusarium Pallidoro - A New Biofertilizer PDFDocument8 pagesFusarium Pallidoro - A New Biofertilizer PDFPaulino Alfonso SuarezPas encore d'évaluation
- Aspergillus Niger Was Used For Cellulase Production in Submerged (SMF) and Solid State Fermentation (SSF)Document9 pagesAspergillus Niger Was Used For Cellulase Production in Submerged (SMF) and Solid State Fermentation (SSF)M Syafiq SamadPas encore d'évaluation
- Isolation, Cloning, in Vitro Cultivation and Growth StudiesDocument21 pagesIsolation, Cloning, in Vitro Cultivation and Growth Studiesbijesh babuPas encore d'évaluation
- Enhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank ReactorsDocument6 pagesEnhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank Reactorsazzahra ghina fadillahPas encore d'évaluation
- Article 4 PDFDocument9 pagesArticle 4 PDFJonathan IslasPas encore d'évaluation
- Content and Amino Acid Composition Certain Evaluated For Microbial Protein ProductionDocument5 pagesContent and Amino Acid Composition Certain Evaluated For Microbial Protein ProductionOlasky GamarraPas encore d'évaluation
- Hao. 1998. Microbial Quality, C. Botulinum, Toxin Production, Fresh Cut Cabbage and LettuceDocument6 pagesHao. 1998. Microbial Quality, C. Botulinum, Toxin Production, Fresh Cut Cabbage and LettucealecheverryPas encore d'évaluation
- Degradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusDocument7 pagesDegradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusMuhammad UmerPas encore d'évaluation
- Jurnal Media SingkongDocument8 pagesJurnal Media SingkongSari Budhi Mei;indaPas encore d'évaluation
- Antimicrobial Property of Gliricidia Sepium Plant Extract: Pakistan J. Agric. Res. Vol 24 No.1-4, 2011Document5 pagesAntimicrobial Property of Gliricidia Sepium Plant Extract: Pakistan J. Agric. Res. Vol 24 No.1-4, 2011Bea Dela ZPas encore d'évaluation
- ArtigoDocument6 pagesArtigoFelipe GomidePas encore d'évaluation
- Production of α-amylase using new strain of Bacillus polymyxa isolated from sweet potatoDocument7 pagesProduction of α-amylase using new strain of Bacillus polymyxa isolated from sweet potatoInternational Organization of Scientific Research (IOSR)Pas encore d'évaluation
- Submerged Culture of Phellinus Linteus - mb-36-178 PDFDocument5 pagesSubmerged Culture of Phellinus Linteus - mb-36-178 PDFAmnuay YeangvorakulPas encore d'évaluation
- Estrategia para La Mejora de La Producción de Prodigiosina Por Un Mutante Serratia Marcescens A Través de La Fermentación Por Lotes de FedDocument4 pagesEstrategia para La Mejora de La Producción de Prodigiosina Por Un Mutante Serratia Marcescens A Través de La Fermentación Por Lotes de FedAndrea Fonnegra ValenciaPas encore d'évaluation
- Biosynthesis and Properties of Extracellular Amylase by Encapsulation Bifidobatrium Bifidum in Batch CultureDocument8 pagesBiosynthesis and Properties of Extracellular Amylase by Encapsulation Bifidobatrium Bifidum in Batch Cultureray m deraniaPas encore d'évaluation
- 1293 FullDocument6 pages1293 FullAndréRochaPas encore d'évaluation
- 9 AltintasDocument8 pages9 AltintasValentina InonizedPas encore d'évaluation
- Effect of Re-Ensiling On The Quality of Sorghum Silage: Acidipropionici, Lactobacillus Plantarum, RelocationDocument8 pagesEffect of Re-Ensiling On The Quality of Sorghum Silage: Acidipropionici, Lactobacillus Plantarum, RelocationDeise Dalazen CastagnaraPas encore d'évaluation
- El Uso de Pectinasa para Tratamiendo de Jugo de Manzana y Mora AzulDocument7 pagesEl Uso de Pectinasa para Tratamiendo de Jugo de Manzana y Mora AzulLaura RDPas encore d'évaluation
- AdesinaDocument11 pagesAdesinaambo wellangPas encore d'évaluation
- Applied and Environmental Microbiology-1986-Bissonnette-838.full - 2020-12-17T162319.697Document4 pagesApplied and Environmental Microbiology-1986-Bissonnette-838.full - 2020-12-17T162319.697hmossPas encore d'évaluation
- Res. J. BiotechDocument5 pagesRes. J. BiotechPatricia GravesPas encore d'évaluation
- Journal Type (Thesis)Document12 pagesJournal Type (Thesis)api-19649313Pas encore d'évaluation
- Evaluation of The Growth Performance of Trichoderma: Harzianum (Rifai.) On Different Culture MediaDocument7 pagesEvaluation of The Growth Performance of Trichoderma: Harzianum (Rifai.) On Different Culture MediakhaerulPas encore d'évaluation
- Growth Response of Activities ofDocument6 pagesGrowth Response of Activities ofSbenmohamedPas encore d'évaluation
- Alkaline ProteaseDocument12 pagesAlkaline ProteaseKaviyarasu VelmuruganPas encore d'évaluation
- 405 FullDocument6 pages405 FullAndréRochaPas encore d'évaluation
- Bjork Et Al 1990 Effects of Growth Rate and Other Factors On Protoplast YieldDocument8 pagesBjork Et Al 1990 Effects of Growth Rate and Other Factors On Protoplast Yieldemilymr3Pas encore d'évaluation
- Wheat-TransformationDocument11 pagesWheat-TransformationVenkat RathinamPas encore d'évaluation
- Production and Optimization of Cellulase From Trichodrma IsolatesDocument12 pagesProduction and Optimization of Cellulase From Trichodrma IsolatesSemAyat GoremsPas encore d'évaluation
- In Vitro Antibacterial and Phytochemical Screening of Jatropha Curcas Seed ExtractDocument5 pagesIn Vitro Antibacterial and Phytochemical Screening of Jatropha Curcas Seed ExtractAdedayo A J AdewumiPas encore d'évaluation
- Journal Type (Thesis)Document12 pagesJournal Type (Thesis)api-19649313Pas encore d'évaluation
- 07 Koomnok Diazotroph Endophytic Bacteria in CultivatedDocument7 pages07 Koomnok Diazotroph Endophytic Bacteria in CultivatedAnnisa DewiPas encore d'évaluation
- Use of Uv-C Treatment For The Inactivation of MicroorganismsDocument6 pagesUse of Uv-C Treatment For The Inactivation of MicroorganismsGonzalo BoninoPas encore d'évaluation
- Bioavailability in of Carbohydrate Material Stored Tropical Fruit SeedsDocument10 pagesBioavailability in of Carbohydrate Material Stored Tropical Fruit SeedsBình MinhPas encore d'évaluation
- Dca3 PDFDocument6 pagesDca3 PDFnagavigneshPas encore d'évaluation
- Efficacy of The Nematophagous Fungus ARF18 in es-ESDocument13 pagesEfficacy of The Nematophagous Fungus ARF18 in es-ESCreyente Del Agua ClaraPas encore d'évaluation
- Comparative Study of Biosurfactants Production Bybacillus Licheniformis and CandidaDocument7 pagesComparative Study of Biosurfactants Production Bybacillus Licheniformis and CandidaCatalinaManjarresPas encore d'évaluation
- The Micro Lora Involved in Aerobic-Thermophilic Sludge StabilizationDocument8 pagesThe Micro Lora Involved in Aerobic-Thermophilic Sludge StabilizationAlex ZomPas encore d'évaluation
- Pichia Anomala J121 During Airtight Storage of Wheat: Nutrient Effects On Biocontrol of Penicillium Roqueforti byDocument5 pagesPichia Anomala J121 During Airtight Storage of Wheat: Nutrient Effects On Biocontrol of Penicillium Roqueforti byNurul FuadyPas encore d'évaluation
- 21-Metabolite+of+P +ostDocument6 pages21-Metabolite+of+P +ostAndrea GPas encore d'évaluation
- Growth Kinetics of Diazotrophic Bacillus Sphaericus UPMB10 Cultured Using Different Types and Concentrations of Carbon and Nitrogen SourcesDocument11 pagesGrowth Kinetics of Diazotrophic Bacillus Sphaericus UPMB10 Cultured Using Different Types and Concentrations of Carbon and Nitrogen SourcesTejas DoshiPas encore d'évaluation
- Fermentation and Recovery of L-Glutamic - Acid - FromDocument8 pagesFermentation and Recovery of L-Glutamic - Acid - FromPrachi BhoirPas encore d'évaluation
- Moisture in Wheat BranDocument7 pagesMoisture in Wheat BranreddygonaPas encore d'évaluation
- CONSERVACION NEISSERIADocument4 pagesCONSERVACION NEISSERIAElizabeth TetelpaPas encore d'évaluation
- Alginate Beads Synthetic Bacteria That Affect: GrowthttDocument10 pagesAlginate Beads Synthetic Bacteria That Affect: GrowthttMelissa BakerPas encore d'évaluation
- Physiological and Microbiological Studies On Production of Alkaline Protease From Locally Isolated Bacillus SubtilisDocument11 pagesPhysiological and Microbiological Studies On Production of Alkaline Protease From Locally Isolated Bacillus SubtilisLaorenciaPas encore d'évaluation
- ChattopadhyayDocument6 pagesChattopadhyayinkaPas encore d'évaluation
- Differential Effects. Differential effects of coinoculations with Pseudomonas jessenii PS06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditionsDocument8 pagesDifferential Effects. Differential effects of coinoculations with Pseudomonas jessenii PS06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditionsemilio_cervantes_7Pas encore d'évaluation
- Development of Callus Colonies From The Isolated Microspore Culture of Capsicum Annuum LDocument6 pagesDevelopment of Callus Colonies From The Isolated Microspore Culture of Capsicum Annuum LAriana ChimiPas encore d'évaluation
- Alkaloids Papaver SomniferumDocument5 pagesAlkaloids Papaver SomniferumZipporah MagantePas encore d'évaluation
- Abadias 2003 Optimization of Growth Conditions of The PostharvestDocument9 pagesAbadias 2003 Optimization of Growth Conditions of The PostharvestFlorencia GorordoPas encore d'évaluation
- Sdarticle 3Document6 pagesSdarticle 3nguyenthao146Pas encore d'évaluation
- Induction and Development of Potato Tubers in A Jar FermentorDocument2 pagesInduction and Development of Potato Tubers in A Jar FermentorRoxana Karen Carhuaz CondoriPas encore d'évaluation
- Differentiation Between Bradyrhizobium Isolates by Colony Type, Exopolysaccharides and Streptomycin ResistanceDocument12 pagesDifferentiation Between Bradyrhizobium Isolates by Colony Type, Exopolysaccharides and Streptomycin ResistanceJuan Carlos Fernandez CernaPas encore d'évaluation
- Phytochemical Screening and in Vitro AntimicrobialDocument4 pagesPhytochemical Screening and in Vitro AntimicrobialArmel Géraldo HOUNDETONPas encore d'évaluation
- Viable Counts Versus The Incidence of Machinery Mold On Processed Fruits and VegetablesDocument4 pagesViable Counts Versus The Incidence of Machinery Mold On Processed Fruits and VegetablesGustavo MoralesPas encore d'évaluation
- Starter Cultures in Food ProductionD'EverandStarter Cultures in Food ProductionBarbara SperanzaPas encore d'évaluation
- Prosopis as a Heat Tolerant Nitrogen Fixing Desert Food Legume: Prospects for Economic Development in Arid LandsD'EverandProsopis as a Heat Tolerant Nitrogen Fixing Desert Food Legume: Prospects for Economic Development in Arid LandsMaria Cecilia PuppoPas encore d'évaluation
- Application of Fluorescent Pseudomonads Inoculant Formulations On Vigna Mungo Through Field TrialDocument5 pagesApplication of Fluorescent Pseudomonads Inoculant Formulations On Vigna Mungo Through Field TrialcarloscarinelliPas encore d'évaluation
- Biofertilizers For SustainabilityDocument26 pagesBiofertilizers For SustainabilitycarloscarinelliPas encore d'évaluation
- Agro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFDocument12 pagesAgro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFcarloscarinelliPas encore d'évaluation
- Recent Advances in Milk Clotting Enzymes: ReviewDocument20 pagesRecent Advances in Milk Clotting Enzymes: ReviewcarloscarinelliPas encore d'évaluation
- Deng2019 PDFDocument8 pagesDeng2019 PDFcarloscarinelliPas encore d'évaluation
- Alternatives to Peat Evaluated for Rhizobia InoculantsDocument9 pagesAlternatives to Peat Evaluated for Rhizobia InoculantscarloscarinelliPas encore d'évaluation
- 2nd Edition UpperIntermediate Tests PDFDocument48 pages2nd Edition UpperIntermediate Tests PDFAna MihaelaPas encore d'évaluation
- Recent Advances in Milk Clotting Enzymes: ReviewDocument20 pagesRecent Advances in Milk Clotting Enzymes: ReviewcarloscarinelliPas encore d'évaluation
- 2nd Edition UpperIntermediate Tests PDFDocument48 pages2nd Edition UpperIntermediate Tests PDFAna MihaelaPas encore d'évaluation
- Tetra Aldose: Aseptic In-Line DosingDocument2 pagesTetra Aldose: Aseptic In-Line DosingcarloscarinelliPas encore d'évaluation
- K Lactis Protein Expression Kit FAQsDocument7 pagesK Lactis Protein Expression Kit FAQscarloscarinelliPas encore d'évaluation
- A Process For Refolding of (Pro-) Chymosin, Comprising Recycling of UreaDocument6 pagesA Process For Refolding of (Pro-) Chymosin, Comprising Recycling of UreacarloscarinelliPas encore d'évaluation
- Biofertilizers For SustainabilityDocument26 pagesBiofertilizers For SustainabilitycarloscarinelliPas encore d'évaluation
- Friuliano 2Document22 pagesFriuliano 2Kiril RavenPas encore d'évaluation
- GE Cross Flow Filtration MethodDocument82 pagesGE Cross Flow Filtration MethodPapagong7077Pas encore d'évaluation
- Growth, Survival and Field Performance of Bradyrhizobial Liquid Inoculant Formulations With Polymeric AdditivesDocument9 pagesGrowth, Survival and Field Performance of Bradyrhizobial Liquid Inoculant Formulations With Polymeric AdditivescarloscarinelliPas encore d'évaluation
- Et Pub 05 PDFDocument36 pagesEt Pub 05 PDFcarloscarinelliPas encore d'évaluation
- Review of antifoam mechanisms in fermentationDocument6 pagesReview of antifoam mechanisms in fermentationcarloscarinelliPas encore d'évaluation
- Survival of Azospirillum Brasilense in Liquid Formulation Amended With Different Chemical Additives PDFDocument4 pagesSurvival of Azospirillum Brasilense in Liquid Formulation Amended With Different Chemical Additives PDFcarloscarinelliPas encore d'évaluation
- Production and Storage of Rhizobium Cell ConcentratesDocument8 pagesProduction and Storage of Rhizobium Cell ConcentratescarloscarinelliPas encore d'évaluation
- Agro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFDocument12 pagesAgro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFcarloscarinelliPas encore d'évaluation
- Review of antifoam mechanisms in fermentationDocument6 pagesReview of antifoam mechanisms in fermentationcarloscarinelliPas encore d'évaluation
- Leghaemoglobin and The SupplyIof O2 To Nitrogen-Fixing Root Nodule Bacteroids Presence of Two Oxidase Systems and ATP Production at Low Free 0, ConcentrationDocument10 pagesLeghaemoglobin and The SupplyIof O2 To Nitrogen-Fixing Root Nodule Bacteroids Presence of Two Oxidase Systems and ATP Production at Low Free 0, ConcentrationcarloscarinelliPas encore d'évaluation
- Inoculant Production and Formulation PDFDocument10 pagesInoculant Production and Formulation PDFcarloscarinelliPas encore d'évaluation
- Production and Storage of Rhizobium Cell ConcentratesDocument8 pagesProduction and Storage of Rhizobium Cell ConcentratescarloscarinelliPas encore d'évaluation
- Biofertilizers For SustainabilityDocument26 pagesBiofertilizers For SustainabilitycarloscarinelliPas encore d'évaluation
- Liquid Inoculant DevelopmentDocument32 pagesLiquid Inoculant DevelopmentcarloscarinelliPas encore d'évaluation
- Agro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFDocument12 pagesAgro Industrial Waste Materials and Wastewater Sludge For Rhizobial Inoculant Production A Review PDFcarloscarinelliPas encore d'évaluation
- Changes in Rennet During StorageDocument8 pagesChanges in Rennet During StoragecarloscarinelliPas encore d'évaluation
- Christian EthicsDocument36 pagesChristian EthicsJoseph FordPas encore d'évaluation
- Alexix Carol Mysticism of Cell CultureDocument20 pagesAlexix Carol Mysticism of Cell CultureARGHA MANNAPas encore d'évaluation
- Membrane Structure and Function in 38 CharactersDocument16 pagesMembrane Structure and Function in 38 CharactersAndleeb ImranPas encore d'évaluation
- BS2001 Tutorial 1Document3 pagesBS2001 Tutorial 1cherlynPas encore d'évaluation
- Pathophysiology of Anemia in CKDDocument6 pagesPathophysiology of Anemia in CKDBazz DantePas encore d'évaluation
- Plant Anatomy: Structure and FunctionsDocument8 pagesPlant Anatomy: Structure and FunctionsWimartama WayanPas encore d'évaluation
- Kanamycin Summary Report 2 Committee Veterinary Medicinal Products - enDocument9 pagesKanamycin Summary Report 2 Committee Veterinary Medicinal Products - enilhamPas encore d'évaluation
- Julian Huxley and The Continuity of Eugenics in Twentieth-Century Britain'Document13 pagesJulian Huxley and The Continuity of Eugenics in Twentieth-Century Britain'Natalina Dass Tera Lodato100% (1)
- ICSE Class 10 Biology Previous Year Question Paper 2012Document10 pagesICSE Class 10 Biology Previous Year Question Paper 2012Bmt KingPas encore d'évaluation
- Parallel Evolution of Cannabinoid Biosynthesis: Nature PlantsDocument31 pagesParallel Evolution of Cannabinoid Biosynthesis: Nature PlantsPaulo EdsonPas encore d'évaluation
- Pathology of Diabetic Ketoacidosis - Romeo Rivera Jr.Document1 pagePathology of Diabetic Ketoacidosis - Romeo Rivera Jr.romeo riveraPas encore d'évaluation
- I.S Transes Quiz 3Document23 pagesI.S Transes Quiz 3Nico LokoPas encore d'évaluation
- Amit Kumar Tyagi PHDDocument40 pagesAmit Kumar Tyagi PHDrohitPas encore d'évaluation
- Sexual Reproduction in PlantsDocument23 pagesSexual Reproduction in PlantsStephen AreriPas encore d'évaluation
- Gastrointestinal Masterclass Session 1Document38 pagesGastrointestinal Masterclass Session 1expandPas encore d'évaluation
- The Weekly Time Table For Year I Semester II of 2016 - 2017 FinalDocument26 pagesThe Weekly Time Table For Year I Semester II of 2016 - 2017 FinalsoePas encore d'évaluation
- Biology 1 - 12 - Q2 - M6Document17 pagesBiology 1 - 12 - Q2 - M6Ken KanekiPas encore d'évaluation
- Histologi Usus Ikan NilaDocument14 pagesHistologi Usus Ikan NilaOnnaSafuraPas encore d'évaluation
- Efficient Ligation of DNADocument21 pagesEfficient Ligation of DNAMariaPas encore d'évaluation
- Identification of Streptococcus Mutans From Human Dental Plaque and Dental Caries Using 16srrna GeneDocument6 pagesIdentification of Streptococcus Mutans From Human Dental Plaque and Dental Caries Using 16srrna GenePeto YahozaPas encore d'évaluation
- Molecular Biology: Louise Kamona Kabwe 1616713: 1001839253:::: / 784199656856434: 100821ASTJ0035Document1 pageMolecular Biology: Louise Kamona Kabwe 1616713: 1001839253:::: / 784199656856434: 100821ASTJ0035Louise KamonaPas encore d'évaluation
- The Rhesus (RH) Blood Group SystemDocument44 pagesThe Rhesus (RH) Blood Group SystemMarl EstradaPas encore d'évaluation
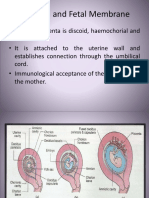
- Placenta and Fetal Membrane-6614Document45 pagesPlacenta and Fetal Membrane-6614Incredible DivinePas encore d'évaluation
- Praktikum FisiologiDocument49 pagesPraktikum FisiologiNurul Fadhilah Taniyo100% (1)
- RFLP Teaching KitDocument5 pagesRFLP Teaching Kitkuldip.biotechPas encore d'évaluation
- Soil Actinomycetes As Potential BiofungicidesDocument5 pagesSoil Actinomycetes As Potential Biofungicideskopeal100% (3)
- SYMPTOMS AND SIGNS OF PLANT DISEASESDocument24 pagesSYMPTOMS AND SIGNS OF PLANT DISEASESNida Ratilla100% (1)
- LAB SG 3 Perchez UPDATES SP18Document4 pagesLAB SG 3 Perchez UPDATES SP18PatriciaPas encore d'évaluation
- Life Science Exit Exam ReviewDocument18 pagesLife Science Exit Exam ReviewJonn EngueroPas encore d'évaluation
- Gizmo Handout Paramecium HomeostasisDocument4 pagesGizmo Handout Paramecium HomeostasisJimena Somoano0% (2)