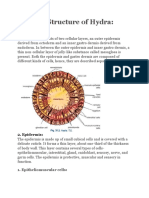
Vous aimerez peut-être aussi
- CoelenterataDocument27 pagesCoelenterataMia LuvIcecream Part IIPas encore d'évaluation
- CNIDARIADocument14 pagesCNIDARIAVanessa DacerPas encore d'évaluation
- HydraDocument6 pagesHydraMishti2100% (2)
- The Phylum Coelenterates: A. General CharacteristicDocument12 pagesThe Phylum Coelenterates: A. General CharacteristicCindy RahmahPas encore d'évaluation
- Histology of Hydra-Types of CellsDocument15 pagesHistology of Hydra-Types of CellsKenneth EbekPas encore d'évaluation
- 3 - Cnidaria and CtenophoraDocument42 pages3 - Cnidaria and CtenophoraDwi HardiyantiPas encore d'évaluation
- Chapter7-Cnidaria Notes2Document8 pagesChapter7-Cnidaria Notes2api-195601294Pas encore d'évaluation
- Exercise 3 Phylum CoelenterataDocument34 pagesExercise 3 Phylum CoelenterataClemence Marie FuentesPas encore d'évaluation
- Primary Dicot Stem and Root in SunflowerDocument10 pagesPrimary Dicot Stem and Root in SunflowerPushkar kumarPas encore d'évaluation
- Nematodes: General CharactersDocument19 pagesNematodes: General CharactersAbdalrhmanPas encore d'évaluation
- Ciliated EpitheliumDocument26 pagesCiliated EpitheliumdeepasanmughamPas encore d'évaluation
- Imad Ullah AssigmentDocument7 pagesImad Ullah AssigmentAsad Ullah KhanPas encore d'évaluation
- Animal Kingdom: Basis of ClassificationDocument15 pagesAnimal Kingdom: Basis of ClassificationChaitali Deshmukh100% (1)
- Aquatic Book PDFDocument105 pagesAquatic Book PDFMohamed TharwatPas encore d'évaluation
- Aurelia MaqniDocument6 pagesAurelia MaqniEhm MelsPas encore d'évaluation
- Anatomy of A Young Dicot RootDocument3 pagesAnatomy of A Young Dicot RootVanessa Leuterio0% (2)
- Phylum PoriferaDocument18 pagesPhylum Poriferagonoles81Pas encore d'évaluation
- Lecture 16 - Annelids & Nematoda: Phylum Annelida Key ConceptDocument19 pagesLecture 16 - Annelids & Nematoda: Phylum Annelida Key ConceptmegsofmemoryPas encore d'évaluation
- Anatomy of Root and StemDocument48 pagesAnatomy of Root and StemLIONS GAMERSPas encore d'évaluation
- Animal Diversity 7th Edition Hickman Solutions ManualDocument8 pagesAnimal Diversity 7th Edition Hickman Solutions Manualkevinwiseeqcydnogzw100% (32)
- HydraDocument3 pagesHydraMagdalenePas encore d'évaluation
- Chapter 10 (Philoid-In)Document8 pagesChapter 10 (Philoid-In)Shrey BhardwajPas encore d'évaluation
- Phylum Cnidaria (Coelenterata)Document112 pagesPhylum Cnidaria (Coelenterata)Silvi valPas encore d'évaluation
- Animal Diversity 7th Edition Hickman Solutions Manual Full Chapter PDFDocument29 pagesAnimal Diversity 7th Edition Hickman Solutions Manual Full Chapter PDFErikaJonestism100% (11)
- ScribdDocument28 pagesScribdGuelan LuarcaPas encore d'évaluation
- EarthwormDocument11 pagesEarthwormFabian GuadarramaPas encore d'évaluation
- CHAPTER 6 Animalia (Invertebrate)Document17 pagesCHAPTER 6 Animalia (Invertebrate)mottong100% (1)
- Culture of Marine PolychaeteDocument7 pagesCulture of Marine PolychaeteyaumilakbarPas encore d'évaluation
- Flatworms, Metazoans and Ribbon WormsDocument59 pagesFlatworms, Metazoans and Ribbon WormsLiz HackettPas encore d'évaluation
- Sbio - Phylum CnidariaDocument36 pagesSbio - Phylum CnidariammballesterosPas encore d'évaluation
- HydraDocument5 pagesHydraBio SciencesPas encore d'évaluation
- Biology - Study of Living Organisms andDocument45 pagesBiology - Study of Living Organisms andNitika GrewalPas encore d'évaluation
- Obelia: Habitat, Structure and DiagramDocument24 pagesObelia: Habitat, Structure and DiagramSunil BundelaPas encore d'évaluation
- Lecture 5 - Cnidarian and Ctenophores - PPT FinalDocument44 pagesLecture 5 - Cnidarian and Ctenophores - PPT FinalSambili TonnyPas encore d'évaluation
- Sponges and Cnidarians 4Document13 pagesSponges and Cnidarians 4kingPas encore d'évaluation
- 40 Life Cycle HerdmaniaDocument7 pages40 Life Cycle HerdmaniaAyushiPas encore d'évaluation
- Ch. 15 Nematoda (Web Version)Document35 pagesCh. 15 Nematoda (Web Version)bewildered-escapist-4301Pas encore d'évaluation
- Paramecium: Chandika KojuDocument11 pagesParamecium: Chandika KojuBhuwan GhimirePas encore d'évaluation
- Anatomy & Plant PhysiologyDocument27 pagesAnatomy & Plant Physiologyshree devPas encore d'évaluation
- Practical 1Document19 pagesPractical 1Nick YuPas encore d'évaluation
- Online Lecture 7Document5 pagesOnline Lecture 7Abdul MunimPas encore d'évaluation
- ScyphozaoaDocument1 pageScyphozaoaNuruddin ZailaniPas encore d'évaluation
- Symmetry and Size: Ghilianii, Is Found in South America and Reaches More ThanDocument3 pagesSymmetry and Size: Ghilianii, Is Found in South America and Reaches More ThanaeiaeiuaoPas encore d'évaluation
- Bio 320 Practical 7Document12 pagesBio 320 Practical 7MuhammadAsyraf50% (2)
- Chordates LabDocument11 pagesChordates LabElda Melissa García RetamaPas encore d'évaluation
- IGCSE Biology NotesDocument50 pagesIGCSE Biology NotesPrincess Kim100% (2)
- Survey of The Phyla-AnimaliaDocument70 pagesSurvey of The Phyla-AnimaliaHelen Gail EmbudoPas encore d'évaluation
- Phylum CnidariaDocument3 pagesPhylum CnidariaMA. LYN CASIPEPas encore d'évaluation
- PhylumDocument15 pagesPhylumNuman AshrafPas encore d'évaluation
- Laboratory 3 Cnidaria and CtenophoraDocument13 pagesLaboratory 3 Cnidaria and Ctenophorapokesurfer100% (1)
- LeechDocument9 pagesLeechХристинаГулеваPas encore d'évaluation
- Safari - Feb 21, 2024 at 11:54 AMDocument1 pageSafari - Feb 21, 2024 at 11:54 AMsyansyncPas encore d'évaluation
- Anim PhylDocument85 pagesAnim PhylTouhidulIslamPas encore d'évaluation
- Monocot and Dicot Root PDFDocument19 pagesMonocot and Dicot Root PDFmukeshtadonPas encore d'évaluation
- Polymorphism in CoelenteratesDocument30 pagesPolymorphism in Coelenteratessrisankaraglobalacademy7Pas encore d'évaluation
- The University of Lahore: Registration No: Submitted By: Submitted To: Subject: TitleDocument20 pagesThe University of Lahore: Registration No: Submitted By: Submitted To: Subject: TitleRafya NoorPas encore d'évaluation
- Hssb2303s Ir SectionDocument3 pagesHssb2303s Ir Sectiontompa kunPas encore d'évaluation
- Major V Sem PracticalDocument12 pagesMajor V Sem Practicalmohitpatel18032003Pas encore d'évaluation
- Solventextraction 150817130936 Lva1 App6892Document37 pagesSolventextraction 150817130936 Lva1 App6892Gowtham GupthaPas encore d'évaluation
- Lecture Notes 7a - Hypothesis Testing For A Population MeanDocument40 pagesLecture Notes 7a - Hypothesis Testing For A Population MeanGowtham GupthaPas encore d'évaluation
- COMP6053 Lecture: Hypothesis Testing, T-Tests, P-Values, Type-I and type-II ErrorsDocument48 pagesCOMP6053 Lecture: Hypothesis Testing, T-Tests, P-Values, Type-I and type-II ErrorsGowtham GupthaPas encore d'évaluation
- Bactrialmorphology 131109235439 Phpapp01Document92 pagesBactrialmorphology 131109235439 Phpapp01GOWTHAM GUPTHAPas encore d'évaluation
- Gen Bio 2nd Quarter (Photosynthesis)Document3 pagesGen Bio 2nd Quarter (Photosynthesis)Andrei Joshua AngellanoPas encore d'évaluation
- TREMATODESDocument36 pagesTREMATODESHearts heavy Moms spaghettiPas encore d'évaluation
- Axonal Degeneration: Nerve RegenerationDocument1 pageAxonal Degeneration: Nerve RegenerationYogi drPas encore d'évaluation
- 1 s2.0 S2352873716300166 Main PDFDocument8 pages1 s2.0 S2352873716300166 Main PDFChalisa ThamdamrongPas encore d'évaluation
- Platelet Physiology: Thomas Gremmel, MD Andrew L. Frelinger Iii, PHD Alan D. Michelson, MDDocument14 pagesPlatelet Physiology: Thomas Gremmel, MD Andrew L. Frelinger Iii, PHD Alan D. Michelson, MDRochnald PigaiPas encore d'évaluation
- Chapter 1 Introduction To BiologyDocument30 pagesChapter 1 Introduction To BiologyGevenia Rissie JenosPas encore d'évaluation
- BZ Lab 5Document8 pagesBZ Lab 5Alexa Jean D. HonrejasPas encore d'évaluation
- Metabolomics - WikipediaDocument12 pagesMetabolomics - WikipediaSamPas encore d'évaluation
- Newsletters 1 3Document6 pagesNewsletters 1 3Ramamoorthy Vanamamalai NanguneriPas encore d'évaluation
- Six Types of PhytoremediationDocument5 pagesSix Types of PhytoremediationChan KianPas encore d'évaluation
- Antiviral DrugsDocument25 pagesAntiviral Drugss.k. kubraPas encore d'évaluation
- The Blue People of Troublesome CreekDocument10 pagesThe Blue People of Troublesome Creekdani77660% (1)
- Is Boron Nutritionally RelevantDocument9 pagesIs Boron Nutritionally RelevantMarisol Alzate FontalvoPas encore d'évaluation
- Mazon - Prelims Histology Week 1Document2 pagesMazon - Prelims Histology Week 1Mazon, Dinah Melisse P.Pas encore d'évaluation
- Lecture 3 Innate ImmunityDocument26 pagesLecture 3 Innate ImmunitytimcarasPas encore d'évaluation
- A.C. Zeven J. Waninge Th. Van Hintum S.P. Singh - Phenotypic Variation in A Core Collection of Common Bean (Phaseolus Vulgaris L.) in The NetherlaDocument14 pagesA.C. Zeven J. Waninge Th. Van Hintum S.P. Singh - Phenotypic Variation in A Core Collection of Common Bean (Phaseolus Vulgaris L.) in The NetherlaNicholas ViniciusPas encore d'évaluation
- Organ Systems 1 NBC ReviewerDocument39 pagesOrgan Systems 1 NBC ReviewerJames ChavezPas encore d'évaluation
- Life Science Final Exam Review-May 2015Document8 pagesLife Science Final Exam Review-May 2015api-230330590Pas encore d'évaluation
- TPHADocument1 pageTPHALidia Narb100% (1)
- Mark Scheme (Results) January 2020: Pearson Edexcel International GCSE in Biology (4BI1) Paper 1BDocument24 pagesMark Scheme (Results) January 2020: Pearson Edexcel International GCSE in Biology (4BI1) Paper 1Bsham67% (3)
- We Got MelaninDocument4 pagesWe Got MelaninLawrence W Spearman57% (7)
- Innovation in Molecular Mechanisms and Tissue EngineeringDocument185 pagesInnovation in Molecular Mechanisms and Tissue EngineeringRenata TereniucPas encore d'évaluation
- Charles Darwin and Natural SelectionDocument10 pagesCharles Darwin and Natural Selectionninaabraham500Pas encore d'évaluation
- JVB 040-2022 - 05Document25 pagesJVB 040-2022 - 05Manuel RuizPas encore d'évaluation
- Nutrition and Wound Healing: Content CreatorsDocument50 pagesNutrition and Wound Healing: Content CreatorsKishan PatelPas encore d'évaluation
- Pharmacology and Its Brief HistoryDocument4 pagesPharmacology and Its Brief HistoryNiala AlmarioPas encore d'évaluation
- Agnarsson and Miller 2008 Acctran Vs DeltranDocument7 pagesAgnarsson and Miller 2008 Acctran Vs DeltranAndrés FrankowPas encore d'évaluation
- Drug Discovery: Finding A Lead: An Introduction To Medicinal Chemistry 3/eDocument26 pagesDrug Discovery: Finding A Lead: An Introduction To Medicinal Chemistry 3/eMima AzrahPas encore d'évaluation
- DNA Origami Activity Guide 1Document4 pagesDNA Origami Activity Guide 1YvonnePas encore d'évaluation