Vous aimerez peut-être aussi
- H1N1Document7 pagesH1N1anon_235643778Pas encore d'évaluation
- H1N1 PDFDocument7 pagesH1N1 PDFanon_235643778Pas encore d'évaluation
- 2015 The Comparison Between Microhematocrit and Automated Methods For Hematocrit DeterminationDocument10 pages2015 The Comparison Between Microhematocrit and Automated Methods For Hematocrit DeterminationAgri Febria SariPas encore d'évaluation
- Thannesberger 2017Document15 pagesThannesberger 2017Денис КрахоткинPas encore d'évaluation
- 140794.3 20210324105625 CoveredDocument16 pages140794.3 20210324105625 CoveredBruno RamírezPas encore d'évaluation
- Nesseria Gonnorhea PaperDocument5 pagesNesseria Gonnorhea PaperHina BatoolPas encore d'évaluation
- Antibody Landscapes After Influenza Virus Infection or VaccinationDocument6 pagesAntibody Landscapes After Influenza Virus Infection or VaccinationManuel Parra CardeñaPas encore d'évaluation
- Gannavaram 2014Document10 pagesGannavaram 2014Shital BishtPas encore d'évaluation
- 2acute Retroviral Syndrome Is Associated With HighDocument10 pages2acute Retroviral Syndrome Is Associated With HighAndrea LunaPas encore d'évaluation
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus InhibitorsDocument15 pagesThe SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus Inhibitorsfaris daboolPas encore d'évaluation
- Comprehensive Serological Profiling of Human Populations Using A Synthetic Human ViromeDocument11 pagesComprehensive Serological Profiling of Human Populations Using A Synthetic Human ViromeembasPas encore d'évaluation
- HHS Public Access: Immune Activation With HIV Vaccines: Implications of The Adenovirus Vector ExperienceDocument6 pagesHHS Public Access: Immune Activation With HIV Vaccines: Implications of The Adenovirus Vector Experiencecomta ccopoPas encore d'évaluation
- Research Paper On h1n1 VirusDocument5 pagesResearch Paper On h1n1 Viruszijkchbkf100% (1)
- The Evolutionary Dynamics of Influenza A Virus Within and Between Human HostsDocument53 pagesThe Evolutionary Dynamics of Influenza A Virus Within and Between Human Hostsaleisha97Pas encore d'évaluation
- Application of Molecular Biology-Based MethodsDocument6 pagesApplication of Molecular Biology-Based MethodsSophie KimPas encore d'évaluation
- Vire MiaDocument8 pagesVire MiaAnderson Rodríguez GuzmanPas encore d'évaluation
- h5n1 Research PapersDocument5 pagesh5n1 Research Papersafeaaawci100% (1)
- A Global Resource For Genomic Predictions of Antimicrobial Resistance and Surveillance of Salmonella Typhi at PathogenwatchDocument12 pagesA Global Resource For Genomic Predictions of Antimicrobial Resistance and Surveillance of Salmonella Typhi at PathogenwatchOkeke UcheePas encore d'évaluation
- VaccineDocument7 pagesVaccineRong LiuPas encore d'évaluation
- Study On Invasive Fungal Infections in Immunocompr PDFDocument7 pagesStudy On Invasive Fungal Infections in Immunocompr PDFMelody PardilloPas encore d'évaluation
- Study On Invasive Fungal Infections in Immunocompr PDFDocument7 pagesStudy On Invasive Fungal Infections in Immunocompr PDFMelody PardilloPas encore d'évaluation
- ASMNGS 2018 AbstractsDocument187 pagesASMNGS 2018 AbstractsturnersdPas encore d'évaluation
- Deyno 2017Document15 pagesDeyno 2017Esteban CastilloPas encore d'évaluation
- Laboratory-Based Versus Population-Based Surveilance of Antimicrobial Resistance To Inform Empirical Treatment For Suspectetd Urinary Tract Infection in IndonesiaDocument10 pagesLaboratory-Based Versus Population-Based Surveilance of Antimicrobial Resistance To Inform Empirical Treatment For Suspectetd Urinary Tract Infection in IndonesiaDavid NicholasPas encore d'évaluation
- International Journal of Antimicrobial Agents: ReviewDocument9 pagesInternational Journal of Antimicrobial Agents: ReviewThania WazintaPas encore d'évaluation
- Journal of Infection: SciencedirectDocument8 pagesJournal of Infection: SciencedirectKarina Dwi SwastikaPas encore d'évaluation
- Review: BackgroundDocument9 pagesReview: BackgroundMichael BalePas encore d'évaluation
- HCV in MexicoDocument8 pagesHCV in MexicoErwin Chiquete, MD, PhDPas encore d'évaluation
- WhoDocument7 pagesWhoYu YingPas encore d'évaluation
- 1 s2.0 S0264410X21006113 MainDocument10 pages1 s2.0 S0264410X21006113 Mainflor rodriguezPas encore d'évaluation
- Test 11Document14 pagesTest 11CiprianPas encore d'évaluation
- Osong Public Health and Research PerspectivesDocument6 pagesOsong Public Health and Research PerspectivesbrianPas encore d'évaluation
- Journal Ppat 1010612Document33 pagesJournal Ppat 1010612ventigvestigePas encore d'évaluation
- MOT 2020 - Transcriptional Profiles of Adjuvanted Hepatitis B Vaccines Display Variable Interindividual Homogeneity But A Shared Core SignatureDocument14 pagesMOT 2020 - Transcriptional Profiles of Adjuvanted Hepatitis B Vaccines Display Variable Interindividual Homogeneity But A Shared Core SignatureJoão Pedro NunesPas encore d'évaluation
- Herpes Simplex Virus Research PaperDocument9 pagesHerpes Simplex Virus Research Paperxsykcbikf100% (3)
- Chou Dhuri 2012Document10 pagesChou Dhuri 2012yakoub bensaadaPas encore d'évaluation
- 146S PDFDocument14 pages146S PDFnittaya mettaPas encore d'évaluation
- HIV Evolution and Diversity ART Treat Van Zyl 2018Document12 pagesHIV Evolution and Diversity ART Treat Van Zyl 2018Juan Salazar PajaresPas encore d'évaluation
- Working Paper 3 h5n1Document7 pagesWorking Paper 3 h5n1vijayPas encore d'évaluation
- Simulated Respiratory Secretion For Use in The DevDocument18 pagesSimulated Respiratory Secretion For Use in The DevRudra BhattacharjeePas encore d'évaluation
- Transcriptome Profiling of Antimicrobial Resistance in Pseudomonas AeruginosaDocument12 pagesTranscriptome Profiling of Antimicrobial Resistance in Pseudomonas Aeruginosapauletta15bpPas encore d'évaluation
- Combining Urine Lipoarabinomannan With Antibody Detection As A Simple Non-Sputum-Based Screening Method For HIV-associated TuberculosisDocument16 pagesCombining Urine Lipoarabinomannan With Antibody Detection As A Simple Non-Sputum-Based Screening Method For HIV-associated TuberculosisJoselinePas encore d'évaluation
- 1 s2.0 S0580951719300236 MainDocument23 pages1 s2.0 S0580951719300236 MainJuan Diego Berrio SaavedraPas encore d'évaluation
- PMID 36974854 OpportunisticDocument8 pagesPMID 36974854 Opportunisticajaya basnetPas encore d'évaluation
- Dengue Full Length ReportDocument101 pagesDengue Full Length ReportEndi VillalobosPas encore d'évaluation
- Evidence That Hepatitis C Virus Genome Partly Controls Infection OutcomeDocument15 pagesEvidence That Hepatitis C Virus Genome Partly Controls Infection OutcomeKaritoPas encore d'évaluation
- Literature Review On Dengue VirusDocument7 pagesLiterature Review On Dengue Virusaflskeqjr100% (1)
- Original Article: Magnitude of Cryptococcal Antigenemia 369Document6 pagesOriginal Article: Magnitude of Cryptococcal Antigenemia 369MegbaruPas encore d'évaluation
- Some Immunological Parameters of Viral Pediatric PneumoniaDocument8 pagesSome Immunological Parameters of Viral Pediatric PneumoniaTJPRC PublicationsPas encore d'évaluation
- FullDocument28 pagesFullPooya AavaniPas encore d'évaluation
- Vaccine InformaticsDocument10 pagesVaccine InformaticsSumathiPas encore d'évaluation
- Towards An Epitope-Based Human Vaccine For Influenza: Tamar Ben-Yedidia Ruth ArnonDocument7 pagesTowards An Epitope-Based Human Vaccine For Influenza: Tamar Ben-Yedidia Ruth ArnonPCRMANPas encore d'évaluation
- 1 s2.0 S1672022915001126 MainDocument8 pages1 s2.0 S1672022915001126 MainRitika PathakPas encore d'évaluation
- Research Paper On The Flu VirusDocument5 pagesResearch Paper On The Flu Viruswyyzcaulg100% (1)
- WHO Manual On Animal Diagnosis and SurveillanceDocument105 pagesWHO Manual On Animal Diagnosis and SurveillanceTalita Batista100% (1)
- Oxford University Press Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Clinical Infectious DiseasesDocument13 pagesOxford University Press Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Clinical Infectious DiseasesPisi MicaPas encore d'évaluation
- Diagnostic Virology: Special Section: Medical MicrobiologyDocument13 pagesDiagnostic Virology: Special Section: Medical MicrobiologyEl FatihPas encore d'évaluation
- Dokumen - Tips PPT Gagal NapaspptxDocument12 pagesDokumen - Tips PPT Gagal NapaspptxIndra Mahardhika PPas encore d'évaluation
- Management of Infections in the Immunocompromised HostD'EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalPas encore d'évaluation
- The Frequency of Influenza and Bacterial Co-Infection - A SystematDocument12 pagesThe Frequency of Influenza and Bacterial Co-Infection - A Systematrahma_husna_1Pas encore d'évaluation
- 7th Edition NRP Brings Big Changes For NRP Instructors: Instructor UpdateDocument12 pages7th Edition NRP Brings Big Changes For NRP Instructors: Instructor UpdateJoev SaquinPas encore d'évaluation
- DMF 40 199 PDFDocument14 pagesDMF 40 199 PDFsiraj_khan_13Pas encore d'évaluation
- World AIDS Day - December 1, 2019 Status of HIV Case-Based Surveillance Implementation - 39 U.S. PEPFAR-Supported Countries, May-July 2019Document16 pagesWorld AIDS Day - December 1, 2019 Status of HIV Case-Based Surveillance Implementation - 39 U.S. PEPFAR-Supported Countries, May-July 2019worksheetbookPas encore d'évaluation
- Glycerol MsdsDocument6 pagesGlycerol MsdsJX Lim0% (1)
- Glossary Mental Terms-FarsiDocument77 pagesGlossary Mental Terms-FarsiRahatullah AlamyarPas encore d'évaluation
- De La Cruz, Et Al. (2015) Treatment of Children With ADHD and IrritabilityDocument12 pagesDe La Cruz, Et Al. (2015) Treatment of Children With ADHD and Irritabilityjuan100% (1)
- Tonsillectomy and Adenoidectomy - Pediatric Clinics of North AmericaDocument13 pagesTonsillectomy and Adenoidectomy - Pediatric Clinics of North AmericayosefPas encore d'évaluation
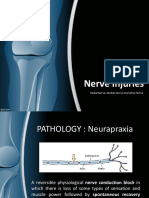
- Nerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveDocument26 pagesNerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveaisiyaahPas encore d'évaluation
- Normative Data For A Spanish Version of The Rey Auditory-Verbal Learning Test in Older PeopleDocument12 pagesNormative Data For A Spanish Version of The Rey Auditory-Verbal Learning Test in Older PeopleeastareaPas encore d'évaluation
- Mental Health Awareness SpeechDocument2 pagesMental Health Awareness SpeechThea Angela Longino95% (20)
- Behaviorism in Daily LifeDocument8 pagesBehaviorism in Daily LifeMichelleTongPas encore d'évaluation
- Job Safety Analysis Worksheet: JSA JSA Participants PPE Required Tools And/or EquipmentDocument5 pagesJob Safety Analysis Worksheet: JSA JSA Participants PPE Required Tools And/or EquipmentVigiePas encore d'évaluation
- Significance of HACCP and SSOP in Food Processing EstablishmentsDocument7 pagesSignificance of HACCP and SSOP in Food Processing EstablishmentselfiraPas encore d'évaluation
- Synthesis EssayDocument8 pagesSynthesis EssayWardah FarukPas encore d'évaluation
- Attitude of EMPLOYEES in Terms of Compliance of Health and SafetyDocument6 pagesAttitude of EMPLOYEES in Terms of Compliance of Health and SafetyJanice KimPas encore d'évaluation
- Hong Kong Dental Journal - Volume 2 - Number 2 - YuLeungDocument1 pageHong Kong Dental Journal - Volume 2 - Number 2 - YuLeungFebySiampaPas encore d'évaluation
- StoryDocument6 pagesStoryGerard NicholasPas encore d'évaluation
- CH 3 VocabDocument5 pagesCH 3 VocabAbdirahman Abdullahi OmarPas encore d'évaluation
- Maternity Benefit-Employer's ObligationsDocument20 pagesMaternity Benefit-Employer's ObligationsSonika BhatiPas encore d'évaluation
- Erik Erikson N BronfennerDocument4 pagesErik Erikson N BronfennerOng Ai LisPas encore d'évaluation
- Vince Gironda 8x8 RoutineDocument10 pagesVince Gironda 8x8 RoutineCLAVDIVS0% (2)
- Pediatrics-AAP Guideline To SedationDocument18 pagesPediatrics-AAP Guideline To SedationMarcus, RNPas encore d'évaluation
- Annual Report of NHRC (2016-17)Document240 pagesAnnual Report of NHRC (2016-17)Shruti Nagvanshi100% (1)
- Usmart 3200T Plus BrochureDocument4 pagesUsmart 3200T Plus BrochureMPas encore d'évaluation
- WBC DisordersDocument45 pagesWBC DisordersyalahopaPas encore d'évaluation
- Analytical Report: Extra-Curricular Activities Name: Tasnuva Radiah Class: Grade 10 Section: 3 Roll No.: 25Document1 pageAnalytical Report: Extra-Curricular Activities Name: Tasnuva Radiah Class: Grade 10 Section: 3 Roll No.: 25Arif AsgarPas encore d'évaluation
- Murawski 2009Document6 pagesMurawski 2009Sofia Valeria MonrealPas encore d'évaluation
- Monitoring The Performance of DamDocument137 pagesMonitoring The Performance of Damputih_138242459100% (2)
- 2022 Arihant Political Science MCQs Term-1 Sample PapersDocument192 pages2022 Arihant Political Science MCQs Term-1 Sample PapersImran Arshad100% (4)
- A 58 Year Old Client Is Admitted With A Diagnosis of Lung CancerDocument9 pagesA 58 Year Old Client Is Admitted With A Diagnosis of Lung CancerNur SanaaniPas encore d'évaluation