Vous aimerez peut-être aussi
- Jneurosci 0073-14 2014Document15 pagesJneurosci 0073-14 2014mahfuz emonPas encore d'évaluation
- Abnormality of Circadian Rhythm and AutismDocument7 pagesAbnormality of Circadian Rhythm and AutismMelissa RomeroPas encore d'évaluation
- Neuroprotective Dopamine - FullDocument7 pagesNeuroprotective Dopamine - FullmorexyzPas encore d'évaluation
- (Sici) 1096 9861 (20000207) 417:2 181::aid Cne4 3.0.co 2 ADocument14 pages(Sici) 1096 9861 (20000207) 417:2 181::aid Cne4 3.0.co 2 AgslifirsPas encore d'évaluation
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9Pas encore d'évaluation
- Tongcui Jiang-文献阅读Document18 pagesTongcui Jiang-文献阅读fareehakanwar93Pas encore d'évaluation
- Cellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaDocument12 pagesCellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaJose GarciaPas encore d'évaluation
- Margery C. Beinfeld Diana M. KorchakDocument8 pagesMargery C. Beinfeld Diana M. KorchakJose GarciaPas encore d'évaluation
- Vitalis 2005Document9 pagesVitalis 2005Raj PrabuPas encore d'évaluation
- Dopamine and Sleep REMDocument13 pagesDopamine and Sleep REMcalfupanPas encore d'évaluation
- Transcription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisDocument11 pagesTranscription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisJose GarciaPas encore d'évaluation
- Colin-Acetil Transferaza PDFDocument25 pagesColin-Acetil Transferaza PDFMarina AlinaPas encore d'évaluation
- Zns 2449Document14 pagesZns 2449Bogdan A. GireadăPas encore d'évaluation
- Prenatal Exposure To Pesticides Disrupts Testicular Histoarchitecture and Alters Testosterone Levels in Male Caiman Latirostris PDFDocument7 pagesPrenatal Exposure To Pesticides Disrupts Testicular Histoarchitecture and Alters Testosterone Levels in Male Caiman Latirostris PDFRiuin23Pas encore d'évaluation
- Vertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsDocument7 pagesVertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsFractalScribd707Pas encore d'évaluation
- TFG-Related Neurologic Disorders: New Insights Into Relationships Between Endoplasmic Reticulum and NeurodegenerationDocument7 pagesTFG-Related Neurologic Disorders: New Insights Into Relationships Between Endoplasmic Reticulum and NeurodegenerationAlana PachecoPas encore d'évaluation
- TMP 5 CCDocument13 pagesTMP 5 CCFrontiersPas encore d'évaluation
- tmp5C3D TMPDocument10 pagestmp5C3D TMPFrontiersPas encore d'évaluation
- Controle Da VentilaçãoDocument7 pagesControle Da VentilaçãoalinestiPas encore d'évaluation
- Cgireprint 13351035 CK NCKDocument10 pagesCgireprint 13351035 CK NCKlenn chandPas encore d'évaluation
- Effects of Sleep Deprivation On Serum Testosterone Concentrations in The RatDocument6 pagesEffects of Sleep Deprivation On Serum Testosterone Concentrations in The RatRico Alfredo HutabaratPas encore d'évaluation
- AlzehimerDocument13 pagesAlzehimerFrancisco SalgadoPas encore d'évaluation
- Mesopontine Organization of Cholinergic and Catecholaminergic Cell Groups in The Normal and Narcoleptic DogDocument13 pagesMesopontine Organization of Cholinergic and Catecholaminergic Cell Groups in The Normal and Narcoleptic DogAndjela RoganovicPas encore d'évaluation
- Brain, Behavior, and ImmunityDocument9 pagesBrain, Behavior, and ImmunityHelio RochaPas encore d'évaluation
- Control of Dorsal Raphe Serotonergic Neurons by The Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABA, and Glutamate ReceptorsDocument13 pagesControl of Dorsal Raphe Serotonergic Neurons by The Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABA, and Glutamate ReceptorsCortate15gPas encore d'évaluation
- Aierqing Et Al., 2019 Comp Juego BecerrosDocument7 pagesAierqing Et Al., 2019 Comp Juego BecerrosJose Luis Ponce CovarrubiasPas encore d'évaluation
- Neurobiologi PenuaanDocument11 pagesNeurobiologi Penuaanjimmy sullivanPas encore d'évaluation
- Schridde2006 IhDocument13 pagesSchridde2006 Ihiulia andreeaPas encore d'évaluation
- Dis. Model. Mech. 2010 Hortopan 144 8Document5 pagesDis. Model. Mech. 2010 Hortopan 144 8Awanish MishraPas encore d'évaluation
- TMP 6 DAEDocument16 pagesTMP 6 DAEFrontiersPas encore d'évaluation
- Neurogenesis Cerebellum-Feb2019Document23 pagesNeurogenesis Cerebellum-Feb2019Luis Beltran-ParrazalPas encore d'évaluation
- Serotonergic ModulationDocument9 pagesSerotonergic Modulationsilvio da costa guerreiroPas encore d'évaluation
- Zhang2020 InteractionDocument18 pagesZhang2020 InteractionJocilene Dantas Torres NascimentoPas encore d'évaluation
- Afafsaggw EscleDocument7 pagesAfafsaggw EscleStephanie DuncanPas encore d'évaluation
- Hypothalamic Arousal Regions Are Activated During Modafinil-Induced WakefulnessDocument9 pagesHypothalamic Arousal Regions Are Activated During Modafinil-Induced WakefulnessYorka OlavarriaPas encore d'évaluation
- Ovarian Hormones Ameliorate Memory Impairment, Cholinergic Deficit, Neuronal Apoptosis and Astrogliosis in A Rat Model of Alzheimer's DiseaseDocument9 pagesOvarian Hormones Ameliorate Memory Impairment, Cholinergic Deficit, Neuronal Apoptosis and Astrogliosis in A Rat Model of Alzheimer's DiseaseHümay ÜnalPas encore d'évaluation
- Crisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Document12 pagesCrisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Saifuddin HaswarePas encore d'évaluation
- Compared Toxicity of The Potassium Channel Blockers, Apamin and DendrotoxinDocument6 pagesCompared Toxicity of The Potassium Channel Blockers, Apamin and DendrotoxinMarcBosqueAlberichPas encore d'évaluation
- Cfos, Jun y ExploracionDocument12 pagesCfos, Jun y ExploracionLuisa Fernanda Acevedo ReyesPas encore d'évaluation
- 1 s2.0 S0143400411005339 Main PDFDocument8 pages1 s2.0 S0143400411005339 Main PDFMarco Antonio AspronPas encore d'évaluation
- Inferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Document19 pagesInferior Colliculus Transcriptome After Status Epilepticus in The Genetically Audiogenic Seizure-Prone Hamster GASH/Sal Sandra M.Javier HerreroPas encore d'évaluation
- Science 1997Document3 pagesScience 1997demmie1989Pas encore d'évaluation
- ATONIADocument15 pagesATONIAMarsa Ulina SilabanPas encore d'évaluation
- Corticalstriatal Interactions in The Generation of Tic-Like Behaviors After Local Striatal DesinhibitionDocument7 pagesCorticalstriatal Interactions in The Generation of Tic-Like Behaviors After Local Striatal DesinhibitionCornelia ParaipanPas encore d'évaluation
- Rinaldi Et Al 2008 Autism PFC Hyperconnectivity HyperplasticityDocument7 pagesRinaldi Et Al 2008 Autism PFC Hyperconnectivity HyperplasticityJortegloriaPas encore d'évaluation
- Tau Protein in Neurodegenerative Diseases - A Review: E V I E WDocument10 pagesTau Protein in Neurodegenerative Diseases - A Review: E V I E WyusufPas encore d'évaluation
- TMP 8208Document15 pagesTMP 8208FrontiersPas encore d'évaluation
- Bienvenu Et Al JNeurosci 2015Document14 pagesBienvenu Et Al JNeurosci 2015Marco CapognaPas encore d'évaluation
- Chapter 17 - Endocrine Systems - 2020 - The Zebrafish in Biomedical ResearchDocument15 pagesChapter 17 - Endocrine Systems - 2020 - The Zebrafish in Biomedical ResearchNicolas BaronPas encore d'évaluation
- A Negative Regulatory Element Required For Light-Dependent Pinopsin Gene ExpressionDocument7 pagesA Negative Regulatory Element Required For Light-Dependent Pinopsin Gene ExpressionFractalScribd707Pas encore d'évaluation
- Direct Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated MigrationDocument11 pagesDirect Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated Migrationhareb_dPas encore d'évaluation
- Deletion of Asparagine Endopeptidase Reduces Anxiety - and Depressive-Like T Behaviors and Improves Abilities of Spatial Cognition in MiceDocument9 pagesDeletion of Asparagine Endopeptidase Reduces Anxiety - and Depressive-Like T Behaviors and Improves Abilities of Spatial Cognition in MiceHümay ÜnalPas encore d'évaluation
- Suto 2005Document10 pagesSuto 2005AlexandraPas encore d'évaluation
- Fncel 08 00148Document18 pagesFncel 08 00148FabricioPas encore d'évaluation
- 6142 FullDocument12 pages6142 FullAmol MundePas encore d'évaluation
- 3 Muscle Sympatethic InnervationDocument7 pages3 Muscle Sympatethic InnervationGeraldi ContrerasPas encore d'évaluation
- ) Pergamon: NeuroscienceDocument15 pages) Pergamon: NeuroscienceSophiaGuedesPas encore d'évaluation
- Abstracts NewerDocument2 pagesAbstracts Newerapi-19812205Pas encore d'évaluation
- Animal Model of EADocument13 pagesAnimal Model of EAzigurat00Pas encore d'évaluation
- What Is Cloning?: Human Cloning Should Be LegalizedDocument5 pagesWhat Is Cloning?: Human Cloning Should Be LegalizedBass MachinePas encore d'évaluation
- Immunology and Immunochemistry 1-1Document70 pagesImmunology and Immunochemistry 1-1Afram MaxwellPas encore d'évaluation
- Jam 14704Document17 pagesJam 14704Raja ChandraPas encore d'évaluation
- RNA ProcessingDocument258 pagesRNA ProcessingStefan ŽivanovićPas encore d'évaluation
- Owen A O'Connor, Stephen M Ansell, John F Seymo 230518 053705Document514 pagesOwen A O'Connor, Stephen M Ansell, John F Seymo 230518 053705Juan RivasPas encore d'évaluation
- Msds HCL 0,1 NDocument6 pagesMsds HCL 0,1 NRinie Aeryn Nayndine'netzzPas encore d'évaluation
- Biotechnology and The Human GoodDocument225 pagesBiotechnology and The Human GoodIvan Castillo Dcv100% (1)
- IAL Biol WBI05 Jan18 Scientific ArticleDocument8 pagesIAL Biol WBI05 Jan18 Scientific ArticleMoeez AkramPas encore d'évaluation
- Field Trial Evaluating The Efficacy of Porcine Epidemic Diarrhea Vaccine, RNA (Harris Vaccine) in The PhilippinesDocument1 pageField Trial Evaluating The Efficacy of Porcine Epidemic Diarrhea Vaccine, RNA (Harris Vaccine) in The PhilippinesJona MayPas encore d'évaluation
- Equilibrium: Application To Drug Design: Nature BiotechnologyDocument5 pagesEquilibrium: Application To Drug Design: Nature BiotechnologysgybleePas encore d'évaluation
- Inflammatory Bowel DiseaseDocument27 pagesInflammatory Bowel DiseaseMihai VladescuPas encore d'évaluation
- Medical TechnologistDocument2 pagesMedical Technologistapi-329932534Pas encore d'évaluation
- Pathoma ChecklistDocument3 pagesPathoma ChecklistChimpPas encore d'évaluation
- Children With DisabilitiesDocument644 pagesChildren With DisabilitiesNeurodesarrollo Infantil100% (2)
- Cell Organelles Worksheet-PrintDocument2 pagesCell Organelles Worksheet-PrintmuraliMuPas encore d'évaluation
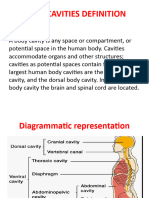
- Body Cavities PPT & Anatomical SystemDocument10 pagesBody Cavities PPT & Anatomical Systemnandhini raguPas encore d'évaluation
- Protein Synthesis: Transcription & TranslationDocument4 pagesProtein Synthesis: Transcription & TranslationtshahriyarPas encore d'évaluation
- Understanding Motor Neuron DiseaseDocument3 pagesUnderstanding Motor Neuron Diseasemasa ePas encore d'évaluation
- NEET GT-5 SolutionsDocument12 pagesNEET GT-5 Solutionsabcxyz7799Pas encore d'évaluation
- University of Cambridge International Examinations General Certificate of Education Ordinary LevelDocument12 pagesUniversity of Cambridge International Examinations General Certificate of Education Ordinary Levelmstudy123456Pas encore d'évaluation
- Pus Poppin FrogsDocument4 pagesPus Poppin FrogsDalia DeebPas encore d'évaluation
- Muscular SystemDocument8 pagesMuscular Systemtheodore_estradaPas encore d'évaluation
- MCQDocument20 pagesMCQanuj sharma100% (2)
- Biochemistry of AgingDocument2 pagesBiochemistry of AgingKaedehara KazuhaPas encore d'évaluation
- 7396 26182 1 PBDocument17 pages7396 26182 1 PBdavidePas encore d'évaluation
- 041 - Endocrinology Physiology) Antidiuretic Hormone (ADH)Document3 pages041 - Endocrinology Physiology) Antidiuretic Hormone (ADH)یوسف رمضان100% (1)
- Nyas12203Becoming Conscious The Science of MindfulnessDocument18 pagesNyas12203Becoming Conscious The Science of MindfulnesscgrossPas encore d'évaluation
- Laboratory Handbook PDFDocument184 pagesLaboratory Handbook PDFReymi Then50% (2)
- Prenatal DevelopmentDocument33 pagesPrenatal DevelopmentDexter Nario0% (1)
- 2022 - Biochemistry - Esha D. Dalvie, Jordan C. Stacy, Keir C. Neuman - Recognition of DNA Supercoil Handedness During Catenation Catalyzed byDocument33 pages2022 - Biochemistry - Esha D. Dalvie, Jordan C. Stacy, Keir C. Neuman - Recognition of DNA Supercoil Handedness During Catenation Catalyzed byjimgogreatPas encore d'évaluation