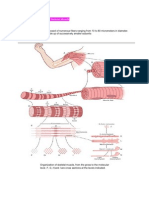
Vous aimerez peut-être aussi
- Sliding Filament Model of ContractionDocument3 pagesSliding Filament Model of ContractionRATU ZULFI AMALIAHPas encore d'évaluation
- Sliding Filament Theory of Muscle ContractionDocument5 pagesSliding Filament Theory of Muscle ContractionRica NorcioPas encore d'évaluation
- Chapter 12 - Muscle PhysiologyDocument21 pagesChapter 12 - Muscle PhysiologyAlec McIntoshPas encore d'évaluation
- He Diagram Below Is A Common One Used To Explain Sliding Filament Theory But Dont Worry About Trying To Understand It All Just YetDocument3 pagesHe Diagram Below Is A Common One Used To Explain Sliding Filament Theory But Dont Worry About Trying To Understand It All Just YetMariam DalilaPas encore d'évaluation
- LECTURE 3. Physiology of Muscles: 1. Functional Anatomy of Skeletal MuscleDocument11 pagesLECTURE 3. Physiology of Muscles: 1. Functional Anatomy of Skeletal MuscleVivek ChaudharyPas encore d'évaluation
- Faal Otot Dan GerakDocument46 pagesFaal Otot Dan GerakElsya Nur FadhillaPas encore d'évaluation
- Cross Bridge CycleDocument14 pagesCross Bridge CycleIdenyi Daniel Ewa EdePas encore d'évaluation
- The Sliding Filament TheoryDocument4 pagesThe Sliding Filament TheoryJinno VeldadPas encore d'évaluation
- 122 3 Long Exam: Muscles and Animal MovementDocument3 pages122 3 Long Exam: Muscles and Animal MovementPia BlancaflorPas encore d'évaluation
- Muscles 1Document25 pagesMuscles 1Mihaela CosteaPas encore d'évaluation
- Muscles 1Document25 pagesMuscles 1Catalina ZamfirPas encore d'évaluation
- L 9 MyosinDocument22 pagesL 9 Myosindiksha singhPas encore d'évaluation
- Muscle Contraction & Sliding Filament TheoryDocument9 pagesMuscle Contraction & Sliding Filament TheoryvishnuPas encore d'évaluation
- Muscles 1Document25 pagesMuscles 1sairash.khan23Pas encore d'évaluation
- Molecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)Document25 pagesMolecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)David HamalalaPas encore d'évaluation
- Actin Myosin CytoskeletonDocument28 pagesActin Myosin Cytoskeletonjesus rajeshdivyaPas encore d'évaluation
- Chapter 5Document25 pagesChapter 5أمال داودPas encore d'évaluation
- Physiology Chap6 (Contraction of Skeletal Muscle)Document5 pagesPhysiology Chap6 (Contraction of Skeletal Muscle)Man DejeloPas encore d'évaluation
- Biochem Report Muscle and CytoskeletonDocument87 pagesBiochem Report Muscle and CytoskeletonKate Lynne CamonayanPas encore d'évaluation
- Skeletal Muscle Contraction and The Motor UnitsDocument4 pagesSkeletal Muscle Contraction and The Motor UnitsAnonymous vnv6QFPas encore d'évaluation
- Costanzo - Fiziologie - Structura SarcomeruluiDocument2 pagesCostanzo - Fiziologie - Structura SarcomeruluiVlad RusuPas encore d'évaluation
- 46 MusclesDocument5 pages46 Musclesuddim004100% (2)
- 11 Skeleton MuscleDocument26 pages11 Skeleton MuscleUSAMA AHMEDPas encore d'évaluation
- Ch-3 Muscle Physiology - Class NotesDocument11 pagesCh-3 Muscle Physiology - Class NotesNarelle IaumaPas encore d'évaluation
- Myosin, Actin & Contraction NotesDocument4 pagesMyosin, Actin & Contraction NotesGhazali BalochPas encore d'évaluation
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120Pas encore d'évaluation
- Miranti Dewi Pramaningtyas Departemen Fisiologi FK UIIDocument37 pagesMiranti Dewi Pramaningtyas Departemen Fisiologi FK UIIFathiZainurohmanPas encore d'évaluation
- Excitable Tissues 1Document8 pagesExcitable Tissues 1gojaisaiah787Pas encore d'évaluation
- Muscoskeletal SystemDocument5 pagesMuscoskeletal SystemJeevikaGoyalPas encore d'évaluation
- Human Anatomy and Physiology - Skeletal MuscleDocument14 pagesHuman Anatomy and Physiology - Skeletal MuscleRAI SUNPas encore d'évaluation
- Physiology of Muscle Contraction and Control of MovementDocument69 pagesPhysiology of Muscle Contraction and Control of MovementMakmunNawil100% (1)
- Mean While,: ("Center" or "Circle") - So, Concentric Things Have A Center in Common. (Document4 pagesMean While,: ("Center" or "Circle") - So, Concentric Things Have A Center in Common. (Karylle SequinaPas encore d'évaluation
- Excitation Contraction CouplingDocument34 pagesExcitation Contraction CouplingRUdraPas encore d'évaluation
- Myofilaments: A) Thick Filaments - The Thick Filament Is Composed of Myosin. TheDocument3 pagesMyofilaments: A) Thick Filaments - The Thick Filament Is Composed of Myosin. TheMichaelJackson Didnt TouchMePas encore d'évaluation
- Muscle Physiology and NeurophysiologyDocument68 pagesMuscle Physiology and NeurophysiologyAlan MagpantayPas encore d'évaluation
- Muscle Tissue Lesson 1Document5 pagesMuscle Tissue Lesson 1Steves LeeboyPas encore d'évaluation
- September 2, 2016 Muscle 1Document37 pagesSeptember 2, 2016 Muscle 1bluemoon4777Pas encore d'évaluation
- Contractile TissuesDocument89 pagesContractile TissuesteePas encore d'évaluation
- Animal Physiology (Autosaved)Document16 pagesAnimal Physiology (Autosaved)Shalinie ChittarasuPas encore d'évaluation
- Sliding Filament Theory Sem 5 AssignmentDocument2 pagesSliding Filament Theory Sem 5 Assignmentparth choudharyPas encore d'évaluation
- Mechanism of Muscle ContractionDocument24 pagesMechanism of Muscle Contractionfisika100% (1)
- Muscles 1Document25 pagesMuscles 1GeorgePas encore d'évaluation
- Muscular SystemDocument10 pagesMuscular SystemOliver MianaPas encore d'évaluation
- Module 10 Kin 267 Muscle TissueDocument4 pagesModule 10 Kin 267 Muscle TissueJ aPas encore d'évaluation
- Biology Lecture Notes: (Stemer'S Guide)Document16 pagesBiology Lecture Notes: (Stemer'S Guide)Ahmed KamelPas encore d'évaluation
- Muscle Physiology NotesDocument20 pagesMuscle Physiology NotesPisiform90100% (1)
- Chapte 3 MuscleDocument128 pagesChapte 3 MuscleErmias HadguPas encore d'évaluation
- Chapter 12 Muscle Physiology - Important Questions: 6. Mechanism of Muscle ContractionDocument7 pagesChapter 12 Muscle Physiology - Important Questions: 6. Mechanism of Muscle ContractionIrum RafeeqPas encore d'évaluation
- Fisiologi MusculoskeletalDocument25 pagesFisiologi MusculoskeletalSuardimanAchoPas encore d'évaluation
- Individual Tissue ConspeqtiDocument16 pagesIndividual Tissue ConspeqtianaPas encore d'évaluation
- Nsca CPTDocument4 pagesNsca CPTRichard Marcus SeigelPas encore d'évaluation
- ST - of Myofilaments..Document10 pagesST - of Myofilaments..Mohammed HamadaPas encore d'évaluation
- Human Physiology 4PHYM001W: Lecture 7 - Muscle Bradley Elliott B.elliott@westminster - Ac.uk @brads - ScienceDocument32 pagesHuman Physiology 4PHYM001W: Lecture 7 - Muscle Bradley Elliott B.elliott@westminster - Ac.uk @brads - ScienceNajma AhmedPas encore d'évaluation
- Introduction To Molecular Motor Proteins: Part 1 Lecture NotesDocument12 pagesIntroduction To Molecular Motor Proteins: Part 1 Lecture Noteskinza888Pas encore d'évaluation
- By Dr.H.Gusbakti, MD, MSC, PKK, AIFM Professor of Physiology University Islamic of North SumateraDocument39 pagesBy Dr.H.Gusbakti, MD, MSC, PKK, AIFM Professor of Physiology University Islamic of North SumateraJennifer WootenPas encore d'évaluation
- Muscle System 2019Document32 pagesMuscle System 2019ZNA EntertainmentPas encore d'évaluation
- Sweeney 2018Document14 pagesSweeney 2018LuisJauweriPas encore d'évaluation
- Lu01 Sliding FilamentDocument4 pagesLu01 Sliding FilamentSuper WarriorPas encore d'évaluation
- Coordination: Physiology of Muscle Contraction and Different Muscle Contraction (Isotonic, Isometric, Isokinetic)Document6 pagesCoordination: Physiology of Muscle Contraction and Different Muscle Contraction (Isotonic, Isometric, Isokinetic)nn1216Pas encore d'évaluation
- Muscular System: The Dog Stole the Professor's NotesD'EverandMuscular System: The Dog Stole the Professor's NotesPas encore d'évaluation
- Week 5 HW Apps Speciation STUDENTSDocument72 pagesWeek 5 HW Apps Speciation STUDENTSSoji Adimula0% (2)
- Solutions To Practice Problems For Genetics, Session 3: PedigreesDocument4 pagesSolutions To Practice Problems For Genetics, Session 3: PedigreesSoji AdimulaPas encore d'évaluation
- LS7B Week 3 Lectures Slides STUDENTSDocument78 pagesLS7B Week 3 Lectures Slides STUDENTSSoji AdimulaPas encore d'évaluation
- Be112atopic15 MusclemechanicsDocument22 pagesBe112atopic15 MusclemechanicsSoji AdimulaPas encore d'évaluation
- Anthony J.F. Griffiths, Susan R. Wessler, Sean B. Carroll, John Doebley Introduction To Genetic Analysis 2010Document862 pagesAnthony J.F. Griffiths, Susan R. Wessler, Sean B. Carroll, John Doebley Introduction To Genetic Analysis 2010Soji Adimula89% (9)
- Chapter 15Document13 pagesChapter 15Soji AdimulaPas encore d'évaluation
- CH 4Document41 pagesCH 4Soji AdimulaPas encore d'évaluation
- Annualcommenceme 1986 NortDocument100 pagesAnnualcommenceme 1986 NortSoji AdimulaPas encore d'évaluation
- Exam3 Practice Problems SolDocument39 pagesExam3 Practice Problems SolSoji AdimulaPas encore d'évaluation
- Today: Eigenstates Vs - Superposition States Measurements: Written HW11 Due On Mon@ NoonDocument15 pagesToday: Eigenstates Vs - Superposition States Measurements: Written HW11 Due On Mon@ NoonSoji AdimulaPas encore d'évaluation
- Etextbook PDF For Study Guide For Pathophysiology The Biological Basis For Disease in Adults and Children 8th EditionDocument62 pagesEtextbook PDF For Study Guide For Pathophysiology The Biological Basis For Disease in Adults and Children 8th Editionreynaldo.bailey262100% (39)
- Activity Modeling Cell RespirationDocument3 pagesActivity Modeling Cell RespirationWendy RatliffPas encore d'évaluation
- All Rights Reserved - Pranav PundarikDocument7 pagesAll Rights Reserved - Pranav PundarikjohnnyPas encore d'évaluation
- Immunology Made Simple AF FinalDocument77 pagesImmunology Made Simple AF FinalStudy EverydayPas encore d'évaluation
- RodakQuestions ComDocument46 pagesRodakQuestions ComMJ ArboledaPas encore d'évaluation
- Sperm Maturation in The Domestic CatDocument11 pagesSperm Maturation in The Domestic CatGeovanni LunaPas encore d'évaluation
- Test Bank For Histology and Cell Biology 2nd Edition Abraham L KierszenbaumDocument5 pagesTest Bank For Histology and Cell Biology 2nd Edition Abraham L KierszenbaumMarta Tronaas100% (38)
- Basic of Proteins - JMCMDocument48 pagesBasic of Proteins - JMCMNeil Vincent De AsisPas encore d'évaluation
- A-Level Biology Revison ChecklistDocument51 pagesA-Level Biology Revison ChecklistImiosa100% (1)
- Modul Hematoimun (Dr. Fakhri) - Immunity To Parasitic InfectionsDocument17 pagesModul Hematoimun (Dr. Fakhri) - Immunity To Parasitic InfectionsAfdaliaPas encore d'évaluation
- Mechanism of Brain Changes in AddictionDocument133 pagesMechanism of Brain Changes in AddictionJared FeldmanPas encore d'évaluation
- Imaging Synapse Structure and FunctionDocument127 pagesImaging Synapse Structure and FunctionFERNANDO ELIAS ALVARENGA CARREROPas encore d'évaluation
- C-Type Lectin Receptor-Induced NF-KB Activation in InnateDocument8 pagesC-Type Lectin Receptor-Induced NF-KB Activation in InnateStefania CristinaPas encore d'évaluation
- How To Construct A Nature Summary Paragraph: Show" or Their Equivalent)Document1 pageHow To Construct A Nature Summary Paragraph: Show" or Their Equivalent)31 Roihan FauziPas encore d'évaluation
- 2 ItemDocument9 pages2 ItemDaniberg RimenisPas encore d'évaluation
- NLR Proteins - Methods and ProtocolsDocument269 pagesNLR Proteins - Methods and ProtocolsG DelisPas encore d'évaluation
- Mitosis MeiosisDocument19 pagesMitosis MeiosisNeo RilloPas encore d'évaluation
- Ieep 105Document7 pagesIeep 105Usha PalPas encore d'évaluation
- 135 - Possible MecanismsDocument19 pages135 - Possible MecanismsEduardo BacelarPas encore d'évaluation
- A Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)Document38 pagesA Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)shamimPas encore d'évaluation
- Biology (Midterm, Hki 2021-2022) 1Document4 pagesBiology (Midterm, Hki 2021-2022) 1Nhi TrầnPas encore d'évaluation
- PhotosynthesisDocument9 pagesPhotosynthesisbok24Pas encore d'évaluation
- MCQ S ZoologyDocument102 pagesMCQ S ZoologySapath GuptaPas encore d'évaluation
- Lesson Plan of Microteaching Direct Instruction Learning ModelDocument22 pagesLesson Plan of Microteaching Direct Instruction Learning ModelFaradillah AsriPas encore d'évaluation
- ExamDocument27 pagesExamKashish HoraPas encore d'évaluation
- PSTcell 3Document2 pagesPSTcell 3Sanja AngPas encore d'évaluation
- Nucleic Acids and Protein SynthesisDocument27 pagesNucleic Acids and Protein SynthesisindraPas encore d'évaluation
- Ws1-Membrane Transport WorksheetDocument3 pagesWs1-Membrane Transport WorksheetTy BakrPas encore d'évaluation
- Protein-Protein Interaction Analysis of Alzheimer's Disease and NAFLD Based On Systems Biology Methods Unhide Common Ancestor PathwaysDocument7 pagesProtein-Protein Interaction Analysis of Alzheimer's Disease and NAFLD Based On Systems Biology Methods Unhide Common Ancestor PathwaysRicardo CastilloPas encore d'évaluation
- Enzymes H2 BIOLOGY NOTESDocument3 pagesEnzymes H2 BIOLOGY NOTESSheila ChaiPas encore d'évaluation